

State-dependent rhythmogenesis and frequency control in a half-center locomotor CPG
- PMID: 28978767
- PMCID: PMC5866471
- DOI: 10.1152/jn.00550.2017
State-dependent rhythmogenesis and frequency control in a half-center locomotor CPG
Erratum in
-
Corrigendum.J Neurophysiol. 2018 Mar 1;119(3):1248-1250. doi: 10.1152/jn.z9k-4583-corr.2018. J Neurophysiol. 2018. PMID: 29537355 Free PMC article. No abstract available.
Abstract
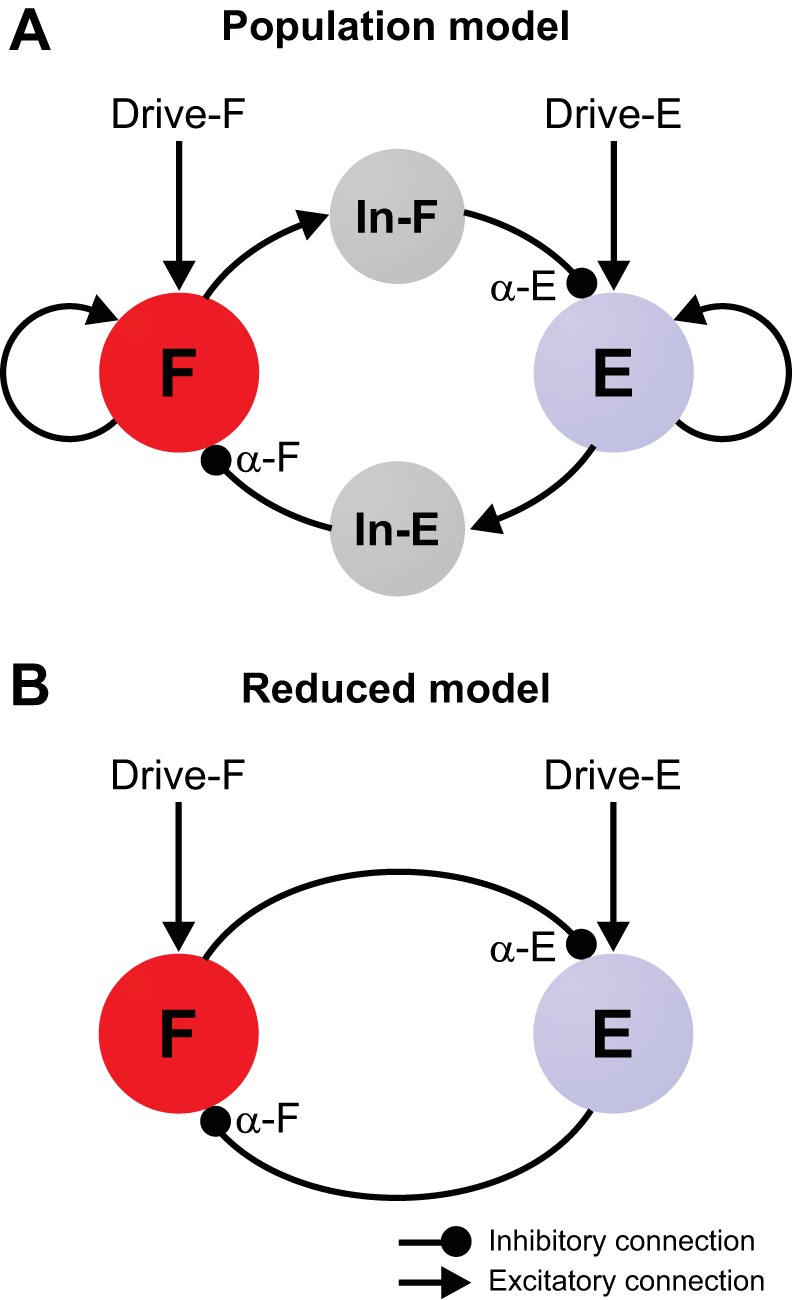
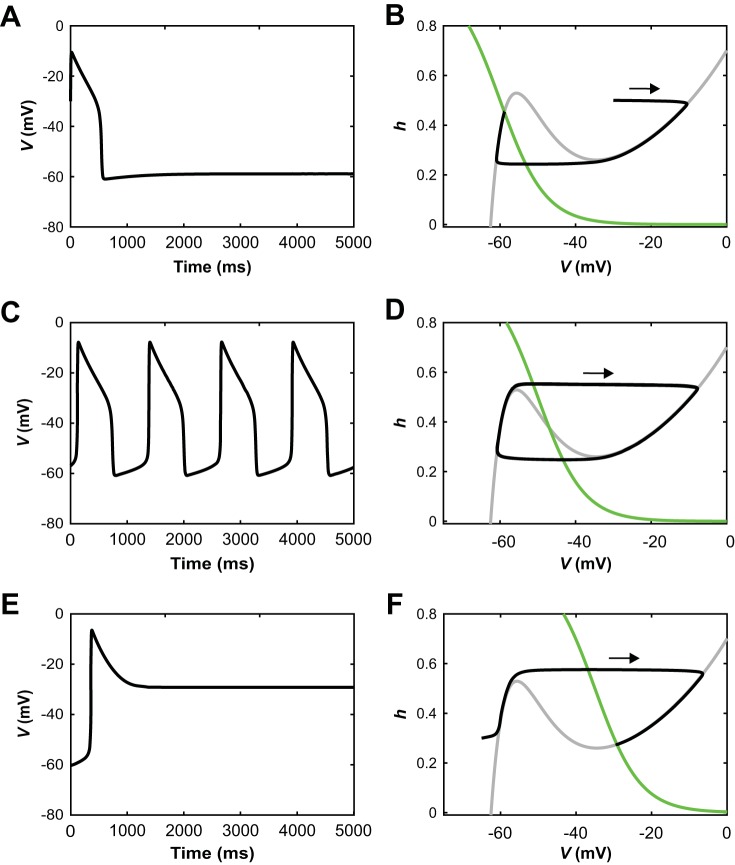
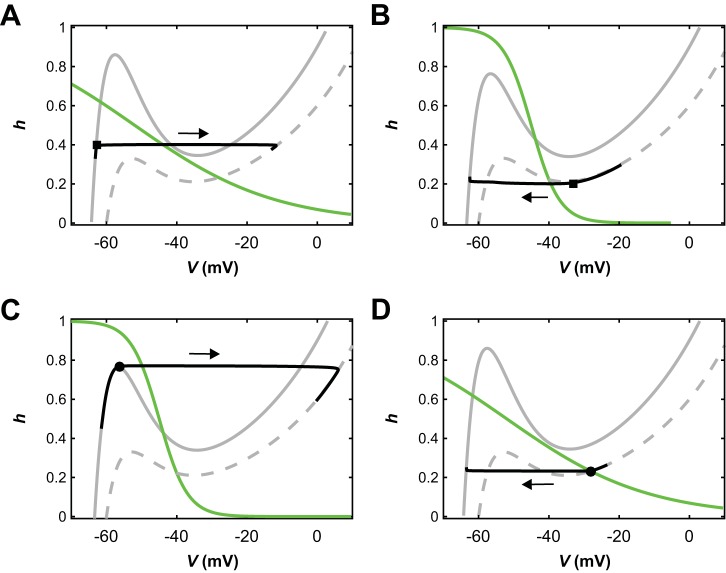
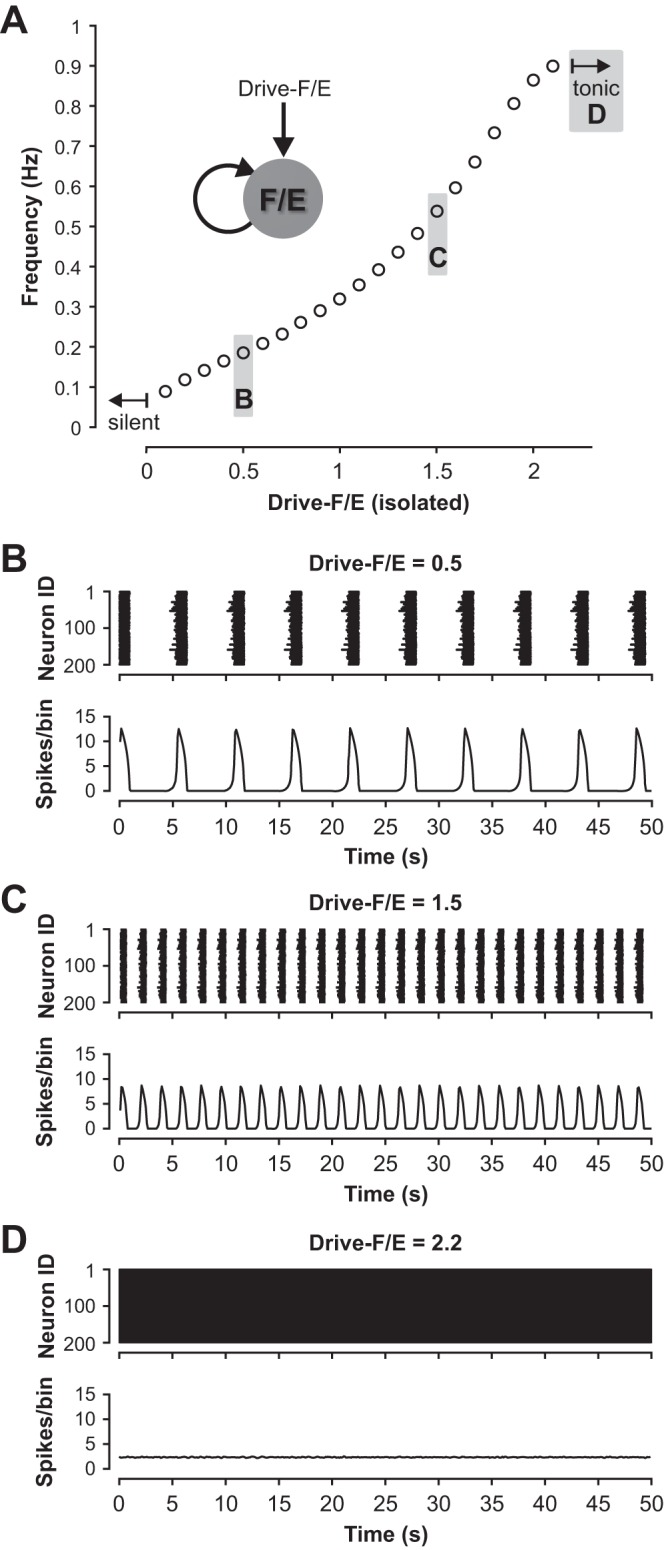
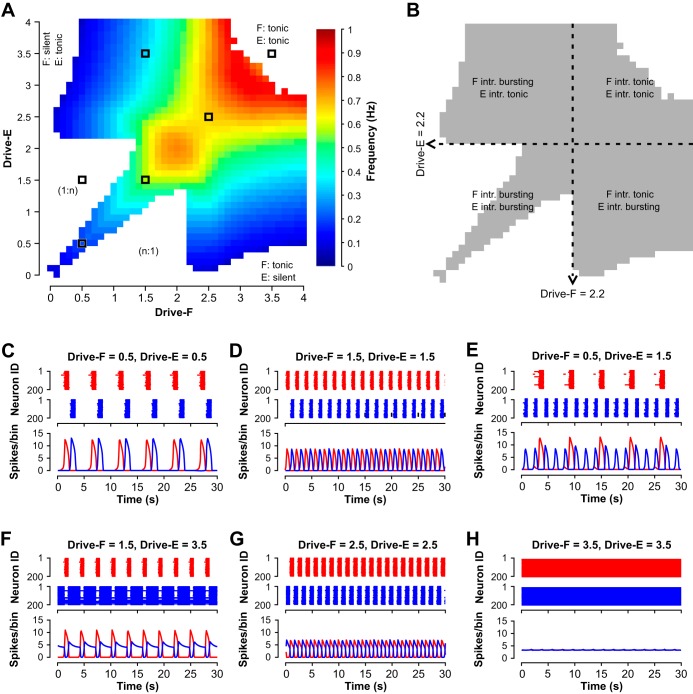
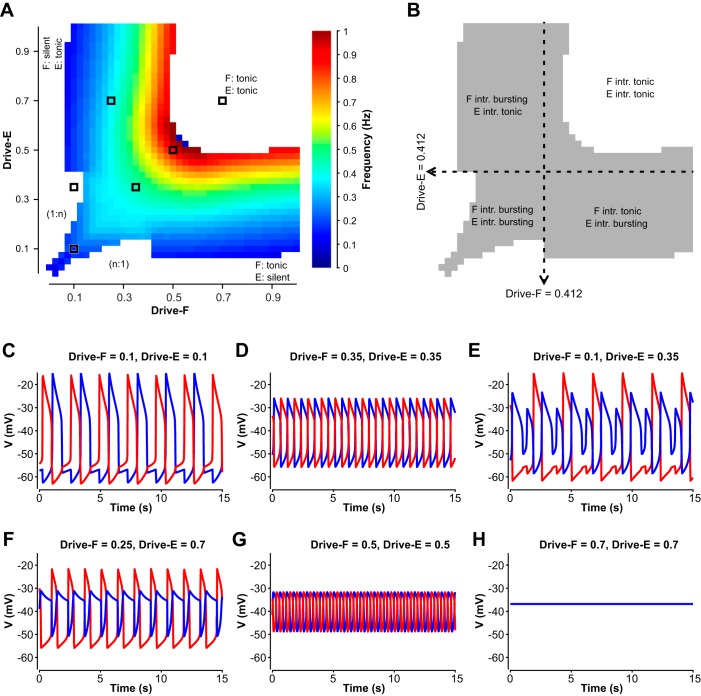
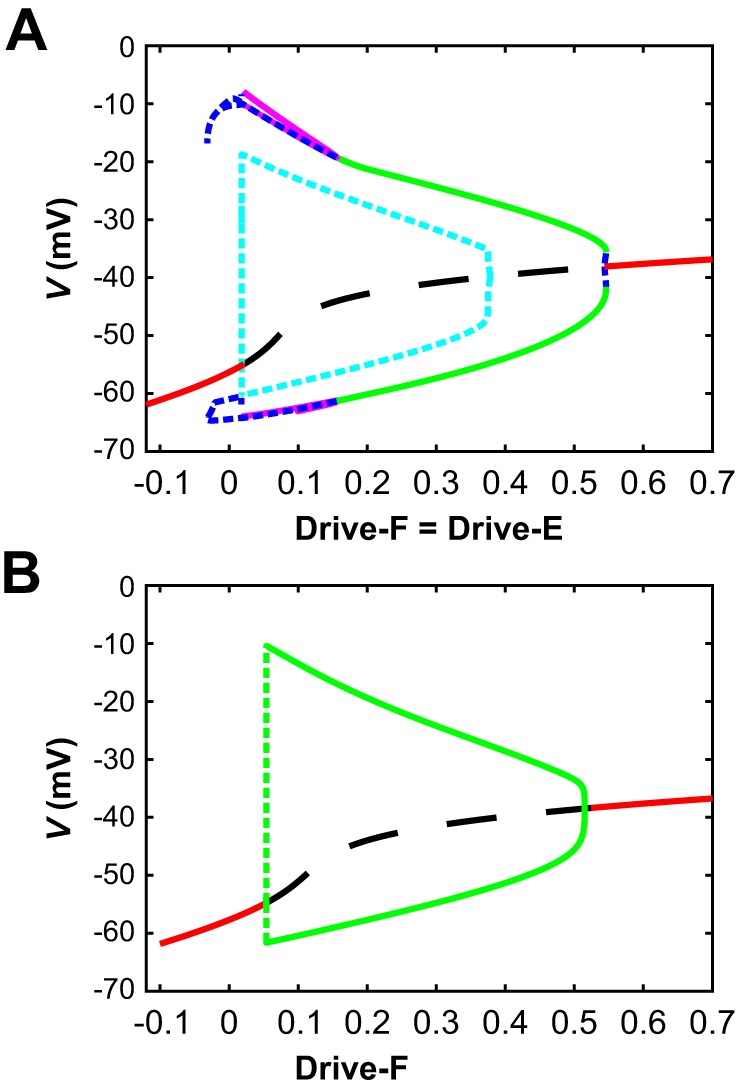
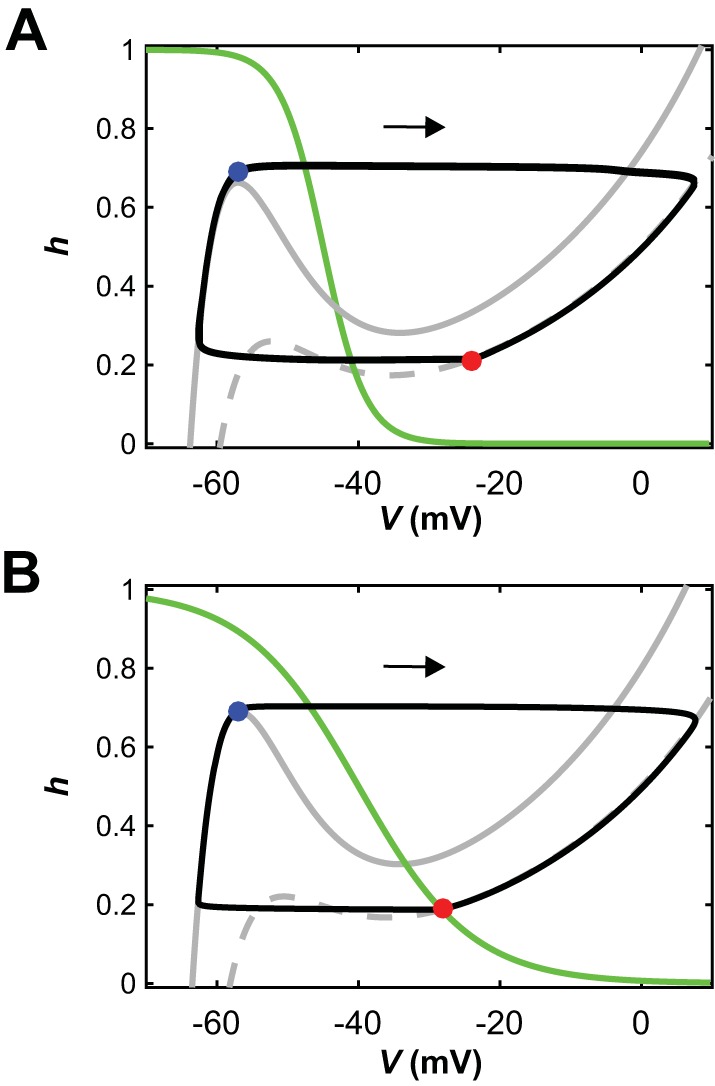
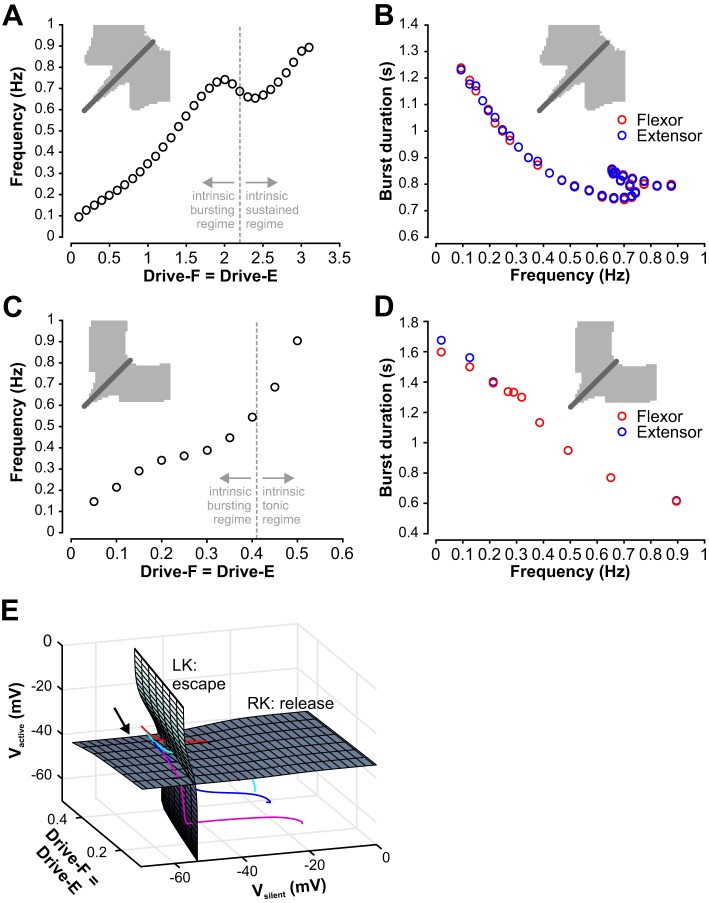
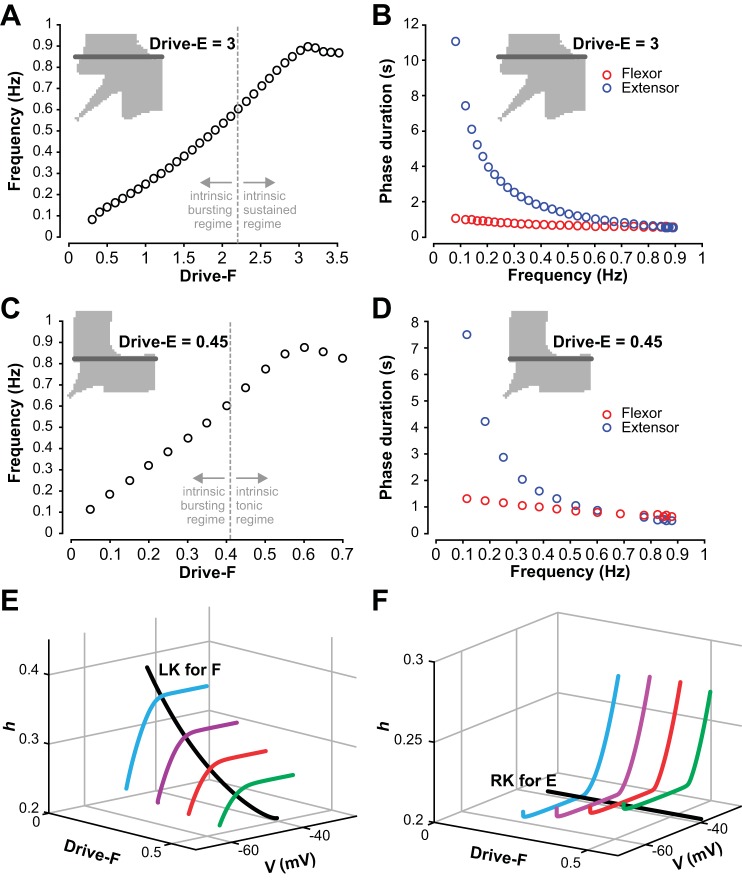
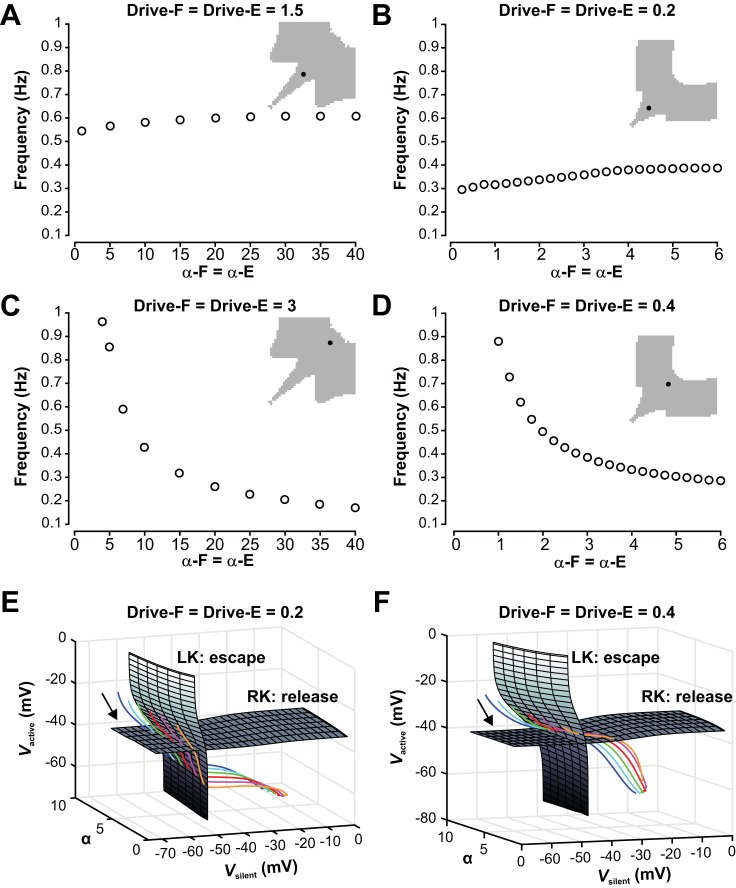
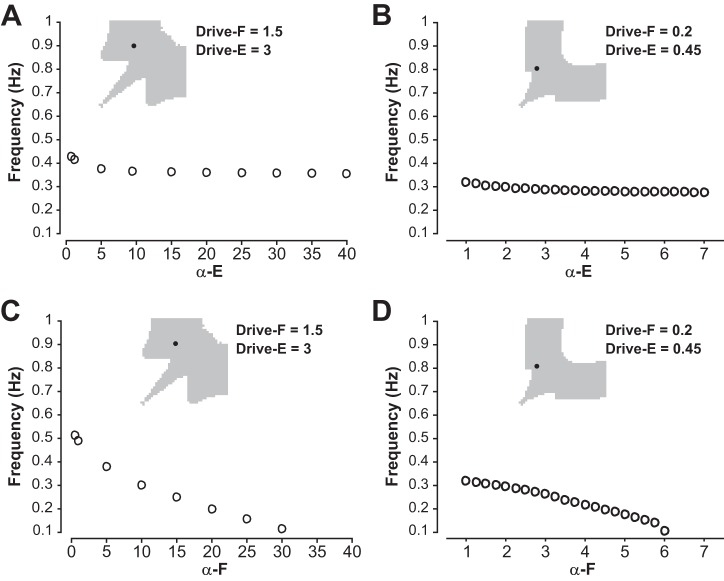
The spinal locomotor central pattern generator (CPG) generates rhythmic activity with alternating flexion and extension phases. This rhythmic pattern is likely to result from inhibitory interactions between neural populations representing flexor and extensor half-centers. However, it is unclear whether the flexor-extensor CPG has a quasi-symmetric organization with both half-centers critically involved in rhythm generation, features an asymmetric organization with flexor-driven rhythmogenesis, or comprises a pair of intrinsically rhythmic half-centers. There are experimental data that support each of the above concepts but appear to be inconsistent with the others. In this theoretical/modeling study, we present and analyze a CPG model architecture that can operate in different regimes consistent with the above three concepts depending on conditions, which are defined by external excitatory drives to CPG half-centers. We show that control of frequency and phase durations within each regime depends on network dynamics, defined by the regime-dependent expression of the half-centers' intrinsic rhythmic capabilities and the operating phase transition mechanisms (escape vs. release). Our study suggests state dependency in locomotor CPG operation and proposes explanations for seemingly contradictory experimental data. NEW & NOTEWORTHY Our theoretical/modeling study focuses on the analysis of locomotor central pattern generators (CPGs) composed of conditionally bursting half-centers coupled with reciprocal inhibition and receiving independent external drives. We show that this CPG framework can operate in several regimes consistent with seemingly contradictory experimental data. In each regime, we study how intrinsic dynamics and phase-switching mechanisms control oscillation frequency and phase durations. Our results provide insights into the organization of spinal circuits controlling locomotion.
Keywords: central pattern generator; computational modeling; flexor-extensor half-center; phase transition mechanisms.
Figures
Similar articles
-
Neuronal activity in the isolated mouse spinal cord during spontaneous deletions in fictive locomotion: insights into locomotor central pattern generator organization.J Physiol. 2012 Oct 1;590(19):4735-59. doi: 10.1113/jphysiol.2012.240895. Epub 2012 Aug 6. J Physiol. 2012. PMID: 22869012 Free PMC article.
-
Asymmetric and transient properties of reciprocal activity of antagonists during the paw-shake response in the cat.PLoS Comput Biol. 2021 Dec 28;17(12):e1009677. doi: 10.1371/journal.pcbi.1009677. eCollection 2021 Dec. PLoS Comput Biol. 2021. PMID: 34962927 Free PMC article.
-
Control of oscillation periods and phase durations in half-center central pattern generators: a comparative mechanistic analysis.J Comput Neurosci. 2009 Aug;27(1):3-36. doi: 10.1007/s10827-008-0124-4. Epub 2009 Jan 6. J Comput Neurosci. 2009. PMID: 19130197 Free PMC article.
-
The CPGs for Limbed Locomotion-Facts and Fiction.Int J Mol Sci. 2021 May 30;22(11):5882. doi: 10.3390/ijms22115882. Int J Mol Sci. 2021. PMID: 34070932 Free PMC article. Review.
-
Modeling the mammalian locomotor CPG: insights from mistakes and perturbations.Prog Brain Res. 2007;165:235-53. doi: 10.1016/S0079-6123(06)65015-2. Prog Brain Res. 2007. PMID: 17925250 Free PMC article. Review.
Cited by
-
Spinal maps of motoneuron activity during human locomotion: neuromechanical considerations.Front Physiol. 2024 Jul 23;15:1389436. doi: 10.3389/fphys.2024.1389436. eCollection 2024. Front Physiol. 2024. PMID: 39108539 Free PMC article. Review.
-
Operation regimes of spinal circuits controlling locomotion and the role of supraspinal drives and sensory feedback.Elife. 2024 Oct 14;13:RP98841. doi: 10.7554/eLife.98841. Elife. 2024. PMID: 39401073 Free PMC article.
-
Robustness of respiratory rhythm generation across dynamic regimes.PLoS Comput Biol. 2019 Jul 30;15(7):e1006860. doi: 10.1371/journal.pcbi.1006860. eCollection 2019 Jul. PLoS Comput Biol. 2019. PMID: 31361738 Free PMC article.
-
A Stochastic Dynamic Operator Framework That Improves the Precision of Analysis and Prediction Relative to the Classical Spike-Triggered Average Method, Extending the Toolkit.eNeuro. 2024 Nov 8;11(11):ENEURO.0512-23.2024. doi: 10.1523/ENEURO.0512-23.2024. Print 2024 Nov. eNeuro. 2024. PMID: 39375031 Free PMC article.
-
Fictive Scratching Patterns in Brain Cortex-Ablated, Midcollicular Decerebrate, and Spinal Cats.Front Neural Circuits. 2020 Feb 27;14:1. doi: 10.3389/fncir.2020.00001. eCollection 2020. Front Neural Circuits. 2020. PMID: 32174815 Free PMC article.
References
-
- Atsuta Y, Garcia-Rill E, Skinner RD. Characteristics of electrically induced locomotion in rat in vitro brain stem-spinal cord preparation. J Neurophysiol 64: 727–735, 1990. - PubMed
-
- Best J, Borisyuk A, Rubin J, Terman D, Wechselberger M. The dynamic range of bursting in a model respiratory pacemaker network. SIAM J Appl Dyn Syst 4: 1107–1139, 2005. doi:10.1137/050625540. - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases