

Emerging Properties and Functional Consequences of Noncoding Transcription
- PMID: 28978770
- PMCID: PMC5629311
- DOI: 10.1534/genetics.117.300095
Emerging Properties and Functional Consequences of Noncoding Transcription
Abstract
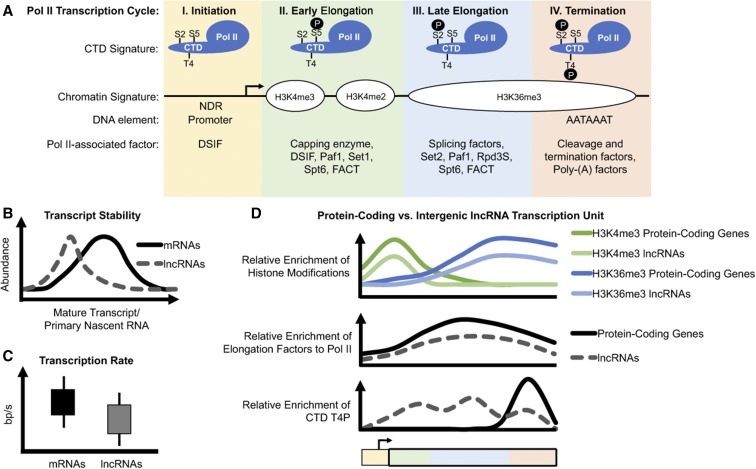
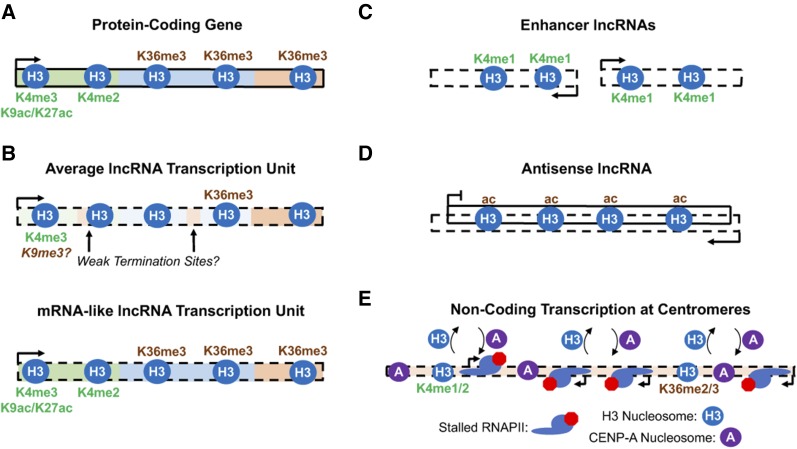
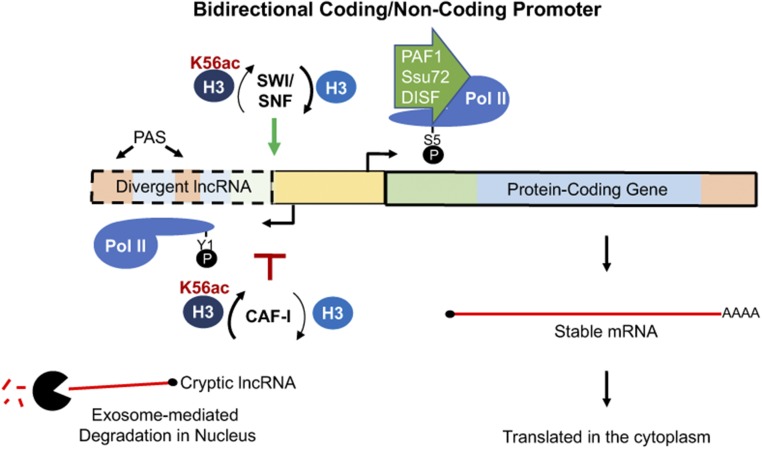
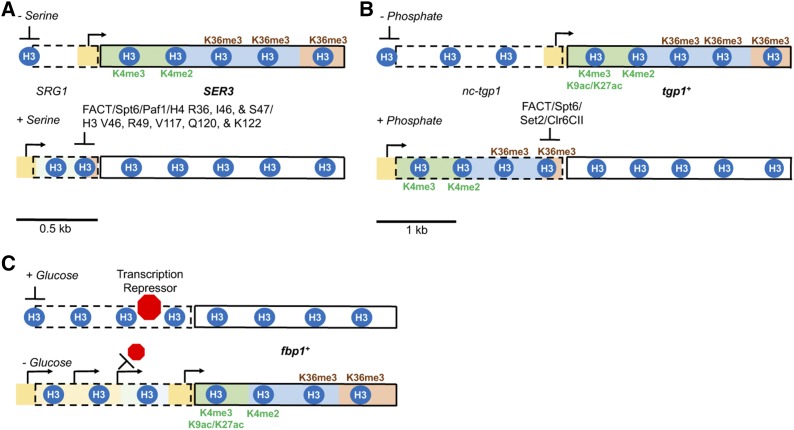
Eukaryotic genomes are rich in transcription units encoding "long noncoding RNAs" (lncRNAs). The purpose of all this transcription is unclear since most lncRNAs are quickly targeted for destruction during synthesis or shortly thereafter. As debates continue over the functional significance of many specific lncRNAs, support grows for the notion that the act of transcription rather than the RNA product itself is functionally important in many cases. Indeed, this alternative mechanism might better explain how low-abundance lncRNAs transcribed from noncoding DNA function in organisms. Here, we highlight some of the recently emerging features that distinguish coding from noncoding transcription and discuss how these differences might have important implications for the functional consequences of noncoding transcription.
Keywords: RNA Polymerase II transcription; chromatin; gene regulation; long noncoding RNA (lncRNA); nascent transcription; noncoding transcription; transcription cycle; transcriptional interference.
Copyright © 2017 by the Genetics Society of America.
Figures
References
-
- Andersson R., Andersen P. R., Valen E., Core L. J., Bornholdt J., et al. , 2014. Nuclear stability and transcriptional directionality separate functionally distinct RNA species. Nat. Commun. 5: 5336. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
