

Allosteric Communication Networks in Proteins Revealed through Pocket Crosstalk Analysis
- PMID: 28979936
- PMCID: PMC5620967
- DOI: 10.1021/acscentsci.7b00211
Allosteric Communication Networks in Proteins Revealed through Pocket Crosstalk Analysis
Abstract
The detection and characterization of binding pockets and allosteric communication in proteins is crucial for studying biological regulation and performing drug design. Nowadays, ever-longer molecular dynamics (MD) simulations are routinely used to investigate the spatiotemporal evolution of proteins. Yet, there is no computational tool that can automatically detect all the pockets and potential allosteric communication networks along these extended MD simulations. Here, we use a novel and fully automated algorithm that examines pocket formation, dynamics, and allosteric communication embedded in microsecond-long MD simulations of three pharmaceutically relevant proteins, namely, PNP, A2A, and Abl kinase. This dynamic analysis uses pocket crosstalk, defined as the temporal exchange of atoms between adjacent pockets, along the MD trajectories as a fingerprint of hidden allosteric communication networks. Importantly, this study indicates that dynamic pocket crosstalk analysis provides new mechanistic understandings on allosteric communication networks, enriching the available experimental data. Thus, our results suggest the prospective use of this unprecedented dynamic analysis to characterize transient binding pockets for structure-based drug design.
Conflict of interest statement
The authors declare the following competing financial interest(s): M.D.V. is scientific advisor, while S.D. and W.R. are shares owners of BiKi Technologies s.r.l., a company that commercializes software solutions for medicinal chemistry including the presented method.
Figures


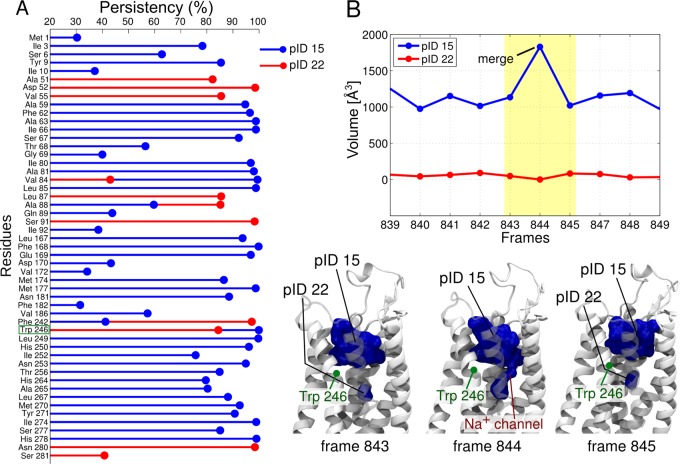
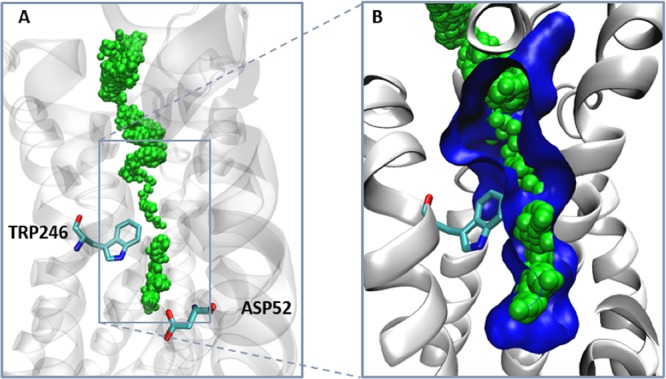
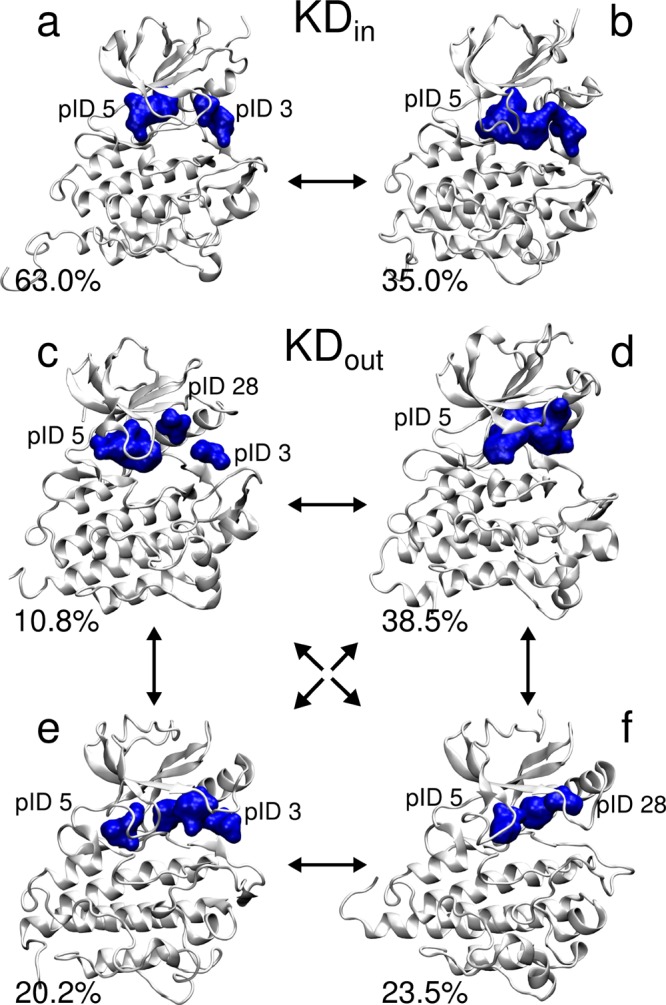

References
-
- Bruce A.; Johnson A.; Lewis J.; Raff M.; Roberts K.; Walter P.. Molecular Biology of the Cell, 4th ed.; Garland Science: New York, 2002.
-
- Spyrakis F.; Benedetti P.; Decherchi S.; Rocchia W.; Cavalli A.; Alcaro S.; Ortuso F.; Baroni M.; Cruciani G. A Pipeline To Enhance Ligand Virtual Screening: Integrating Molecular Dynamics and Fingerprints for Ligand and Proteins. J. Chem. Inf. Model. 2015, 55 (10), 2256–2274. 10.1021/acs.jcim.5b00169. - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
