

Sampling and energy evaluation challenges in ligand binding protein design
- PMID: 28980354
- PMCID: PMC5699494
- DOI: 10.1002/pro.3317
Sampling and energy evaluation challenges in ligand binding protein design
Abstract
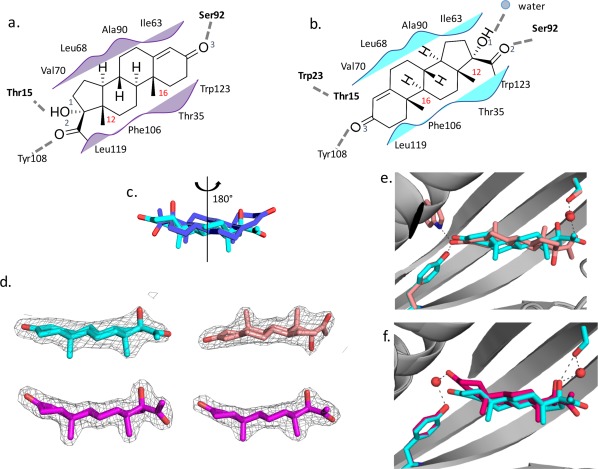
The steroid hormone 17α-hydroxylprogesterone (17-OHP) is a biomarker for congenital adrenal hyperplasia and hence there is considerable interest in development of sensors for this compound. We used computational protein design to generate protein models with binding sites for 17-OHP containing an extended, nonpolar, shape-complementary binding pocket for the four-ring core of the compound, and hydrogen bonding residues at the base of the pocket to interact with carbonyl and hydroxyl groups at the more polar end of the ligand. Eight of 16 designed proteins experimentally tested bind 17-OHP with micromolar affinity. A co-crystal structure of one of the designs revealed that 17-OHP is rotated 180° around a pseudo-two-fold axis in the compound and displays multiple binding modes within the pocket, while still interacting with all of the designed residues in the engineered site. Subsequent rounds of mutagenesis and binding selection improved the ligand affinity to nanomolar range, while appearing to constrain the ligand to a single bound conformation that maintains the same "flipped" orientation relative to the original design. We trace the discrepancy in the design calculations to two sources: first, a failure to model subtle backbone changes which alter the distribution of sidechain rotameric states and second, an underestimation of the energetic cost of desolvating the carbonyl and hydroxyl groups of the ligand. The difference between design model and crystal structure thus arises from both sampling limitations and energy function inaccuracies that are exacerbated by the near two-fold symmetry of the molecule.
Keywords: 17-hydroxylprogesterone; Computational protein design; hydrophobic small molecules; ligand binding design.
© 2017 The Authors Protein Science published by Wiley Periodicals, Inc. on behalf of The Protein Society.
Figures




References
-
- Speiser PW, White PC (2003) Congenital adrenal hyperplasia. N Engl J Med 349:776–788. - PubMed
-
- White PC (2009) Neonatal screening for congenital adrenal hyperplasia. Nat Rev Endocrinol 5:490–498. - PubMed
-
- Griss R, Schena A, Reymond L, Patiny L, Werner D, Tinberg CE, Baker D, Johnsson K (2014) Bioluminescent sensor proteins for point‐of‐care therapeutic drug monitoring. Nat Chem Biol 10:598–603. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
