

Naturally existing isoforms of miR-222 have distinct functions
- PMID: 28981911
- PMCID: PMC5737821
- DOI: 10.1093/nar/gkx788
Naturally existing isoforms of miR-222 have distinct functions
Abstract
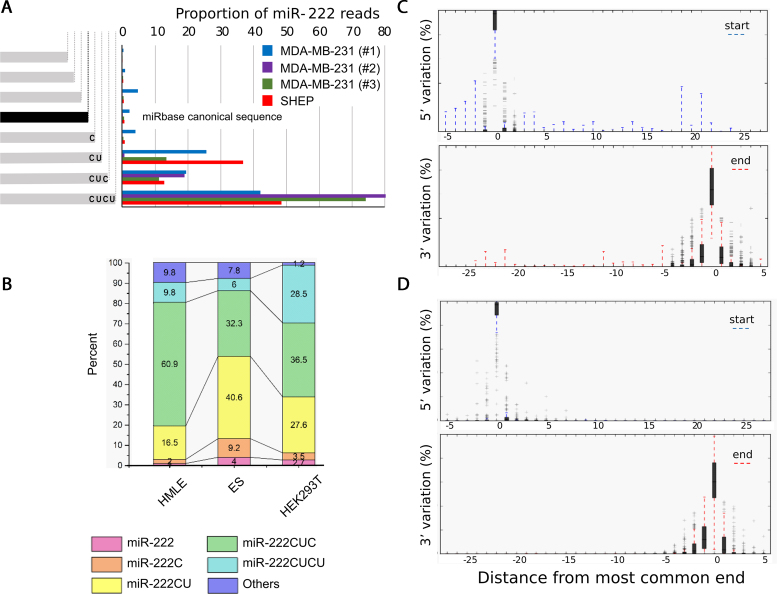
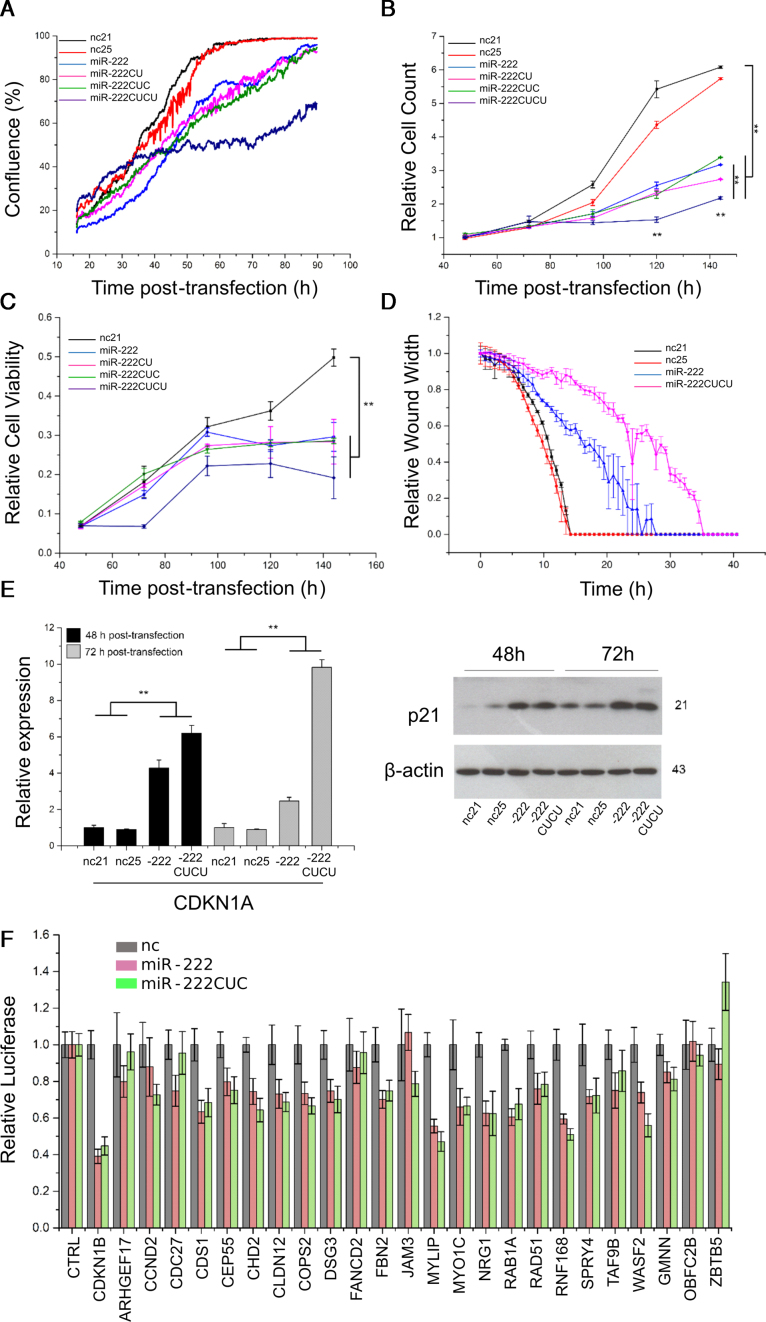
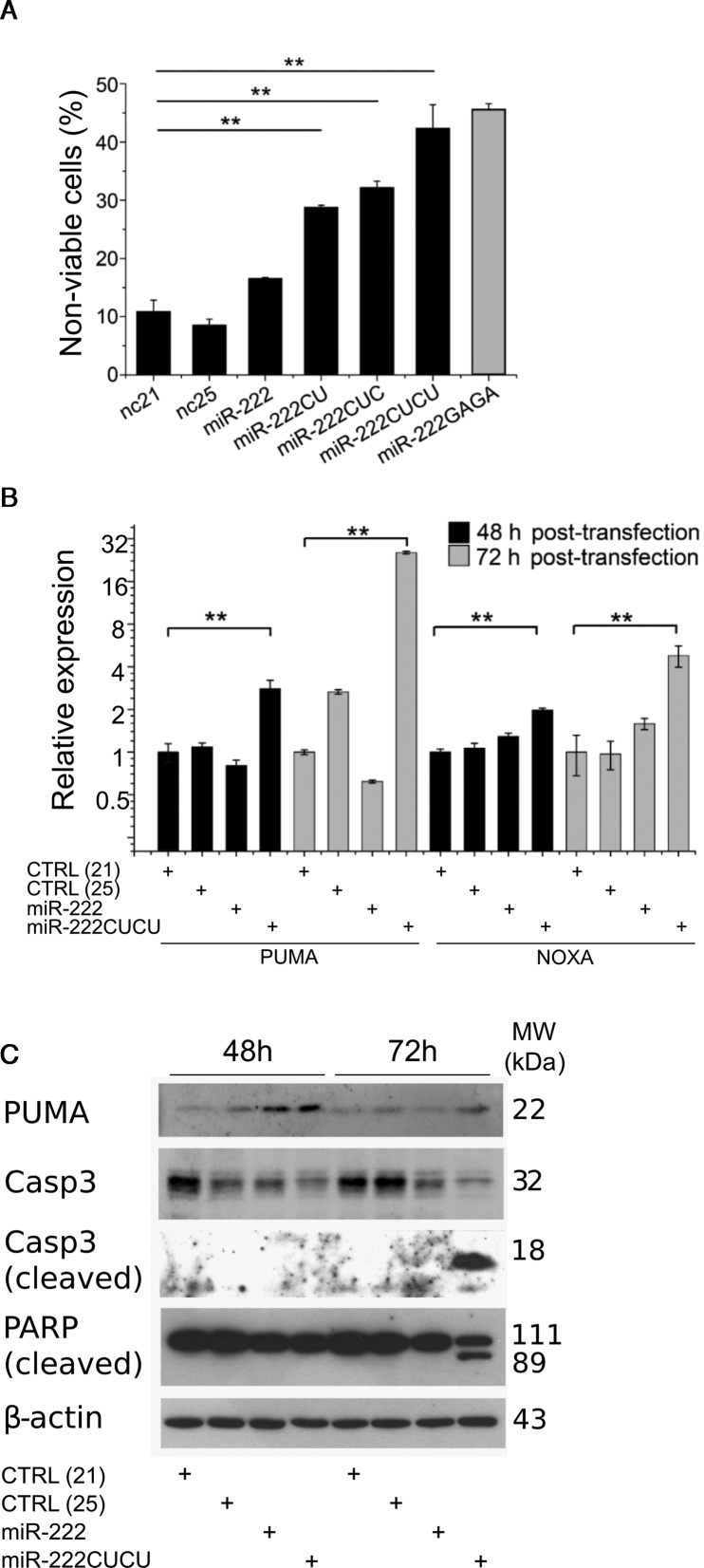
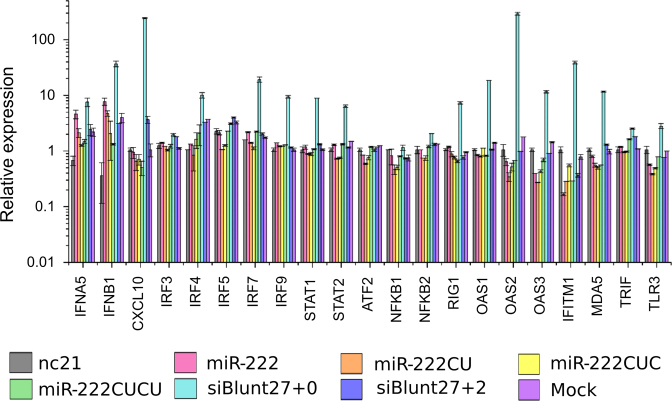
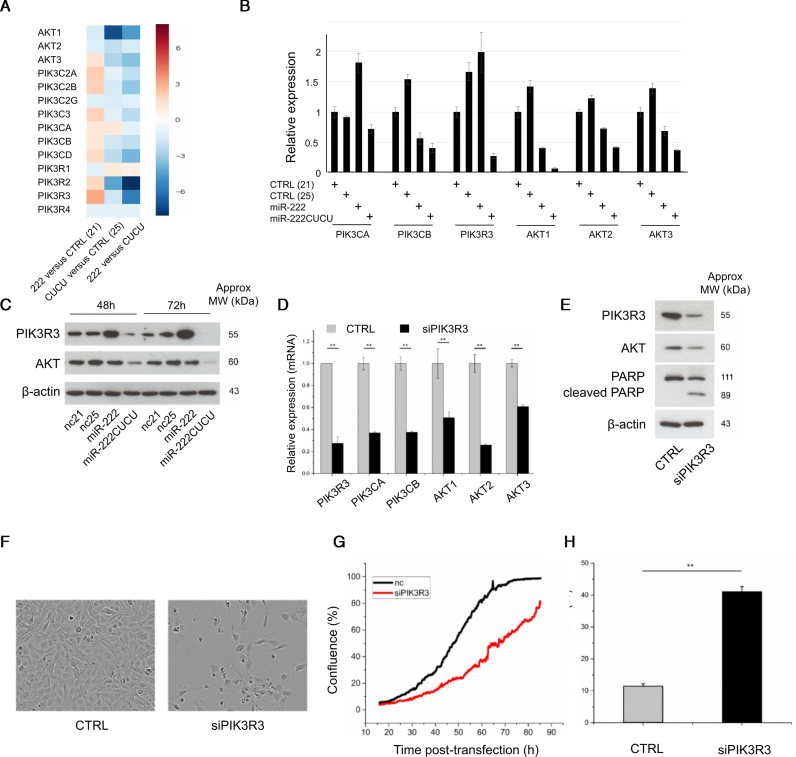
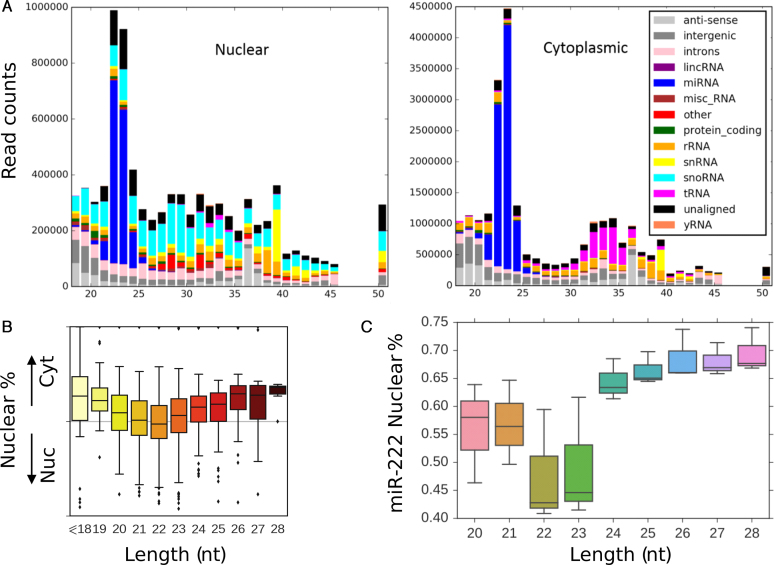
Deep-sequencing reveals extensive variation in the sequence of endogenously expressed microRNAs (termed 'isomiRs') in human cell lines and tissues, especially in relation to the 3' end. From the immunoprecipitation of the microRNA-binding protein Argonaute and the sequencing of associated small RNAs, we observe extensive 3'-isomiR variation, including for miR-222 where the majority of endogenously expressed miR-222 is extended by 1-5 nt compared to the canonical sequence. We demonstrate this 3' heterogeneity has dramatic implications for the phenotype of miR-222 transfected cells, with longer isoforms promoting apoptosis in a size (but not 3' sequence)-dependent manner. The transfection of longer miR-222 isomiRs did not induce an interferon response, but did downregulate the expression of many components of the pro-survival PI3K-AKT pathway including PIK3R3, a regulatory subunit whose knockdown phenocopied the expression of longer 222 isoforms in terms of apoptosis and the inhibition of other PI3K-AKT genes. As this work demonstrates the capacity for 3' isomiRs to mediate differential functions, we contend more attention needs to be given to 3' variance given the prevalence of this class of isomiR.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures

References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
