

Neural Signature of Value-Based Sensorimotor Prioritization in Humans
- PMID: 28982706
- PMCID: PMC5666589
- DOI: 10.1523/JNEUROSCI.1164-17.2017
Neural Signature of Value-Based Sensorimotor Prioritization in Humans
Abstract
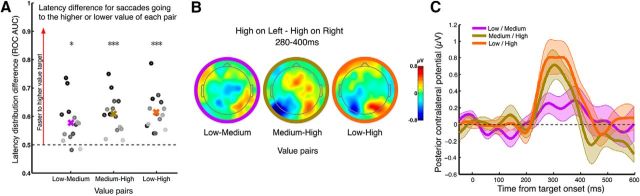
In situations in which impending sensory events demand fast action choices, we must be ready to prioritize higher-value courses of action to avoid missed opportunities. When such a situation first presents itself, stimulus-action contingencies and their relative value must be encoded to establish a value-biased state of preparation for an impending sensorimotor decision. Here, we sought to identify neurophysiological signatures of such processes in the human brain (both female and male). We devised a task requiring fast action choices based on the discrimination of a simple visual cue in which the differently valued sensory alternatives were presented 750-800 ms before as peripheral "targets" that specified the stimulus-action mapping for the upcoming decision. In response to the targets, we identified a discrete, transient, spatially selective signal in the event-related potential (ERP), which scaled with relative value and strongly predicted the degree of behavioral bias in the upcoming decision both across and within subjects. This signal is not compatible with any hitherto known ERP signature of spatial selection and also bears novel distinctions with respect to characterizations of value-sensitive, spatially selective activity found in sensorimotor areas of nonhuman primates. Specifically, a series of follow-up experiments revealed that the signal was reliably invoked regardless of response laterality, response modality, sensory feature, and reward valence. It was absent, however, when the response deadline was relaxed and the strategic need for biasing removed. Therefore, more than passively representing value or salience, the signal appears to play a versatile and active role in adaptive sensorimotor prioritization.SIGNIFICANCE STATEMENT In many situations such as fast-moving sports, we must be ready to act fast in response to sensory events and, in our preparation, prioritize courses of action that lead to greater rewards. Although behavioral effects of value biases in sensorimotor decision making have been widely studied, little is known about the neural processes that set these biases in place beforehand. Here, we report the discovery of a transient, spatially selective neural signal in humans that encodes the relative value of competing decision alternatives and strongly predicts behavioral value biases in decisions made ∼500 ms later. Follow-up manipulations of value differential, reward valence, response modality, sensory features, and time constraints establish that the signal reflects an active, feature- and effector-general preparatory mechanism for value-based prioritization.
Keywords: ERP; decision making; human; sensorimotor; urgency; value bias.
Copyright © 2017 the authors 0270-6474/17/3710725-13$15.00/0.
Figures







References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources