

Capturing the Asc1p/ R eceptor for A ctivated C K inase 1 (RACK1) Microenvironment at the Head Region of the 40S Ribosome with Quantitative BioID in Yeast
- PMID: 28982715
- PMCID: PMC5724181
- DOI: 10.1074/mcp.M116.066654
Capturing the Asc1p/ R eceptor for A ctivated C K inase 1 (RACK1) Microenvironment at the Head Region of the 40S Ribosome with Quantitative BioID in Yeast
Abstract
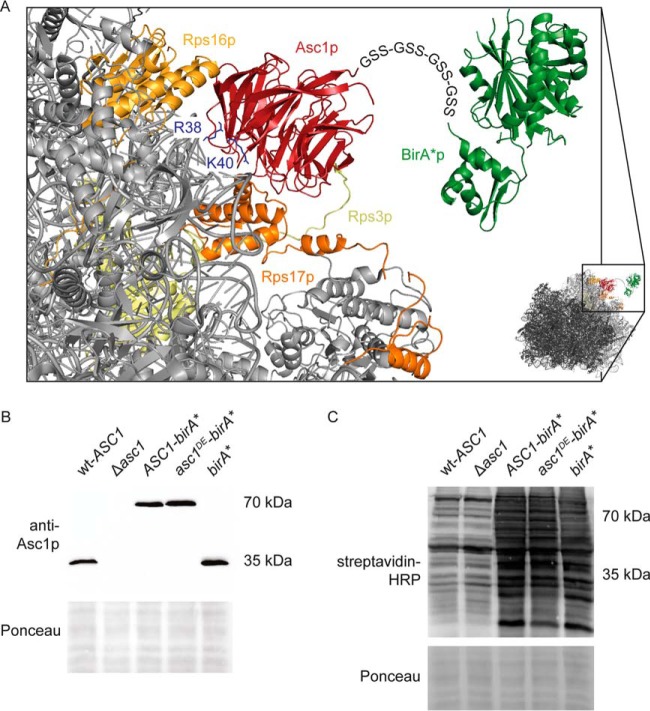
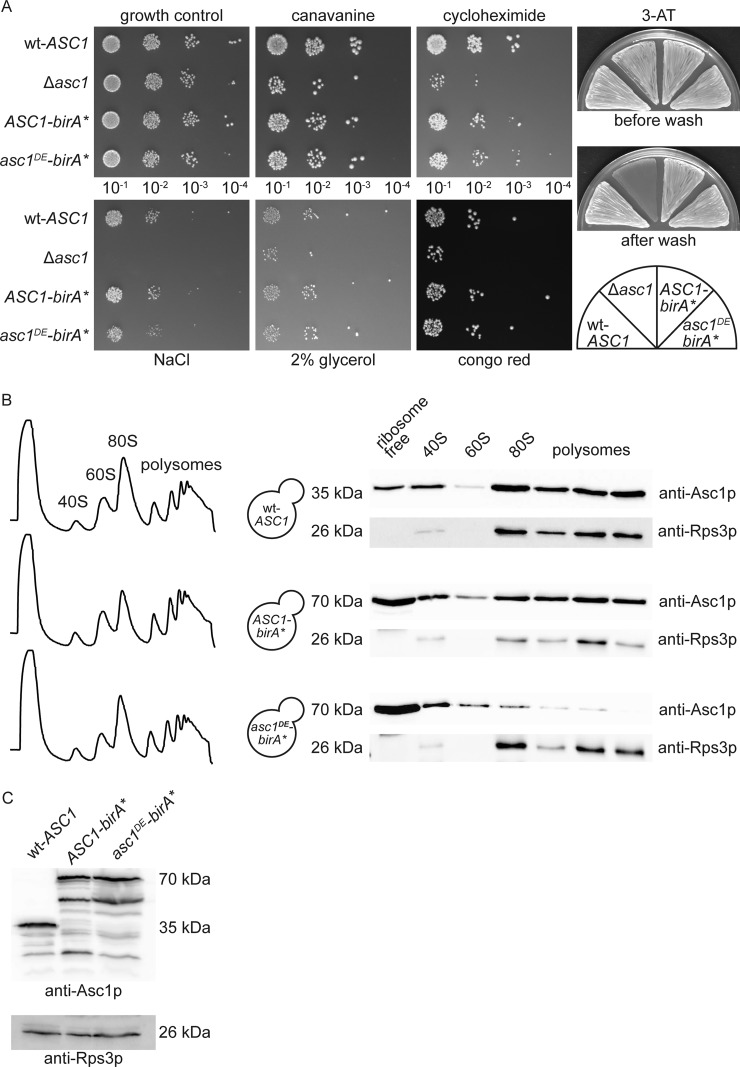
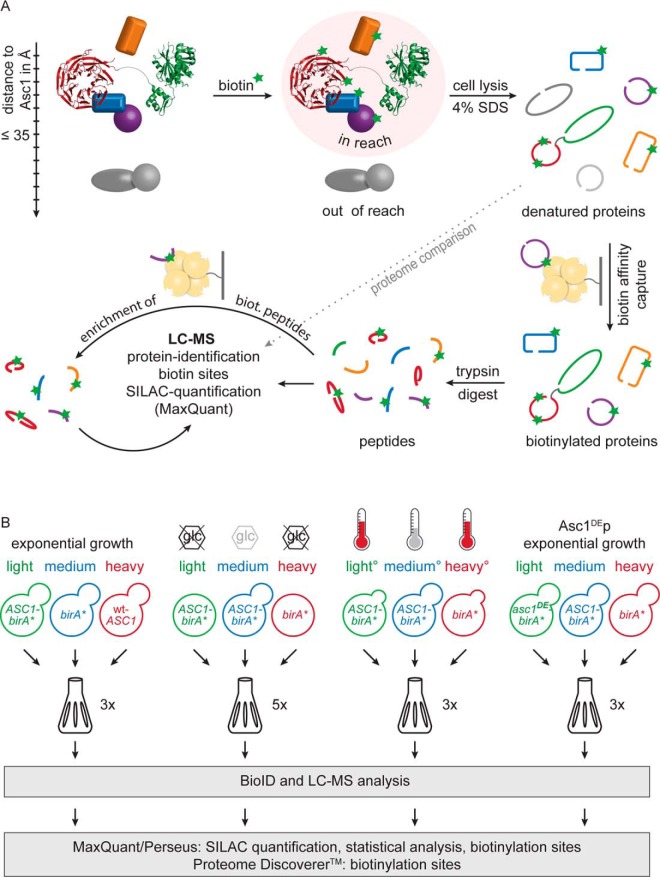
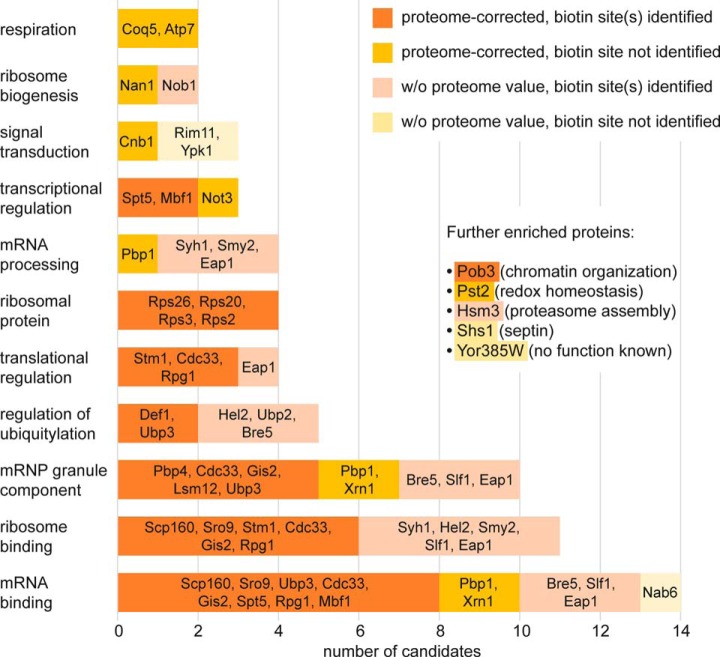
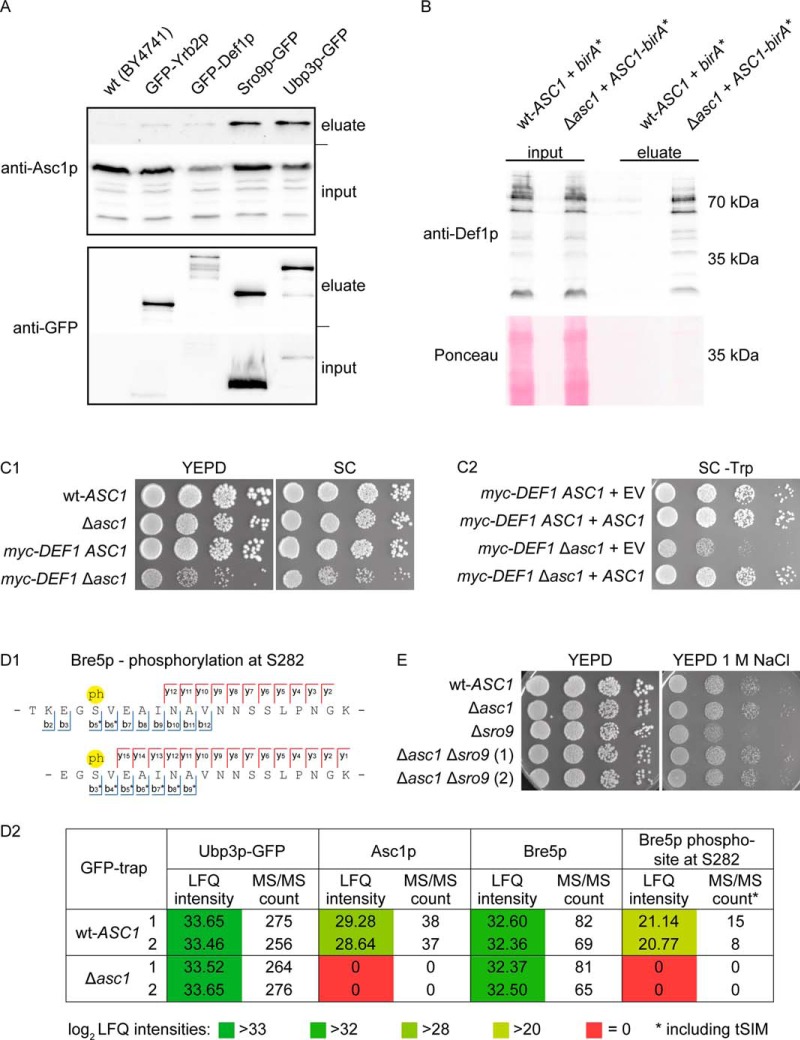
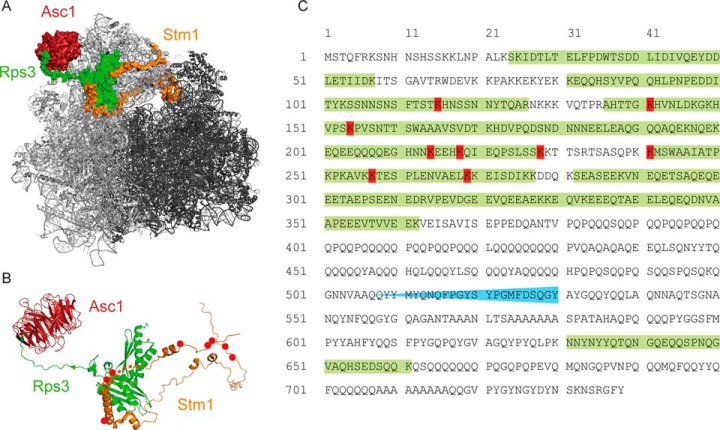
The Asc1 protein of Saccharomyces cerevisiae is a scaffold protein at the head region of ribosomal 40S that links mRNA translation to cellular signaling. In this study, proteins that colocalize with Asc1p were identified with proximity-dependent Biotin IDentification (BioID), an in vivo labeling technique described here for the first time for yeast. Biotinylated Asc1p-birA*-proximal proteins were identified and quantitatively verified against controls applying SILAC and mass spectrometry. The mRNA-binding proteins Sro9p and Gis2p appeared together with Scp160p, each providing ribosomes with nuclear transcripts. The cap-binding protein eIF4E (Cdc33p) and the eIF3/a-subunit (Rpg1p) were identified reflecting the encounter of proteins involved in the initiation of mRNA translation at the head region of ribosomal 40S. Unexpectedly, a protein involved in ribosome preservation (the clamping factor Stm1p), the deubiquitylation complex Ubp3p-Bre5p, the RNA polymerase II degradation factor 1 (Def1p), and transcription factors (Spt5p, Mbf1p) colocalize with Asc1p in exponentially growing cells. For Asc1R38D, K40Ep, a variant considered to be deficient in binding to ribosomes, BioID revealed its predominant ribosome localization. Glucose depletion replaced most of the Asc1p colocalizing proteins for additional ribosomal proteins, suggesting a ribosome aggregation process during early nutrient limitation, possibly concomitant with ribosomal subunit clamping. Overall, the characterization of the Asc1p microenvironment with BioID confirmed and substantiated our recent findings that the β-propeller broadly contributes to signal transduction influencing phosphorylation of colocalizing proteins (e.g. of Bre5p), and by that might affect nuclear gene transcription and the fate of ribosomes.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures

References
-
- Pan C. Q., Sudol M., Sheetz M., and Low B. C. (2012) Modularity and functional plasticity of scaffold proteins as p(l) acemakers in cell signaling. Cell Signal 24, 2143–2165 - PubMed
-
- Vondriska T. M., Pass J. M., and Ping P. (2004) Scaffold proteins and assembly of multiprotein signaling complexes. J. Mol. Cell Cardiol 37, 391–397 - PubMed
-
- Chantrel Y., Gaisne M., Lions C., and Verdière J. (1998) The transcriptional regulator hap1p (cyp1p) is essential for anaerobic or heme-deficient growth of saccharomyces cerevisiae: genetic and molecular characterization of an extragenic suppressor that encodes a wd repeat protein. Genetics 148, 559–569 - PMC - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
