

CAMTA-Mediated Regulation of Salicylic Acid Immunity Pathway Genes in Arabidopsis Exposed to Low Temperature and Pathogen Infection
- PMID: 28982964
- PMCID: PMC5774559
- DOI: 10.1105/tpc.16.00865
CAMTA-Mediated Regulation of Salicylic Acid Immunity Pathway Genes in Arabidopsis Exposed to Low Temperature and Pathogen Infection
Abstract
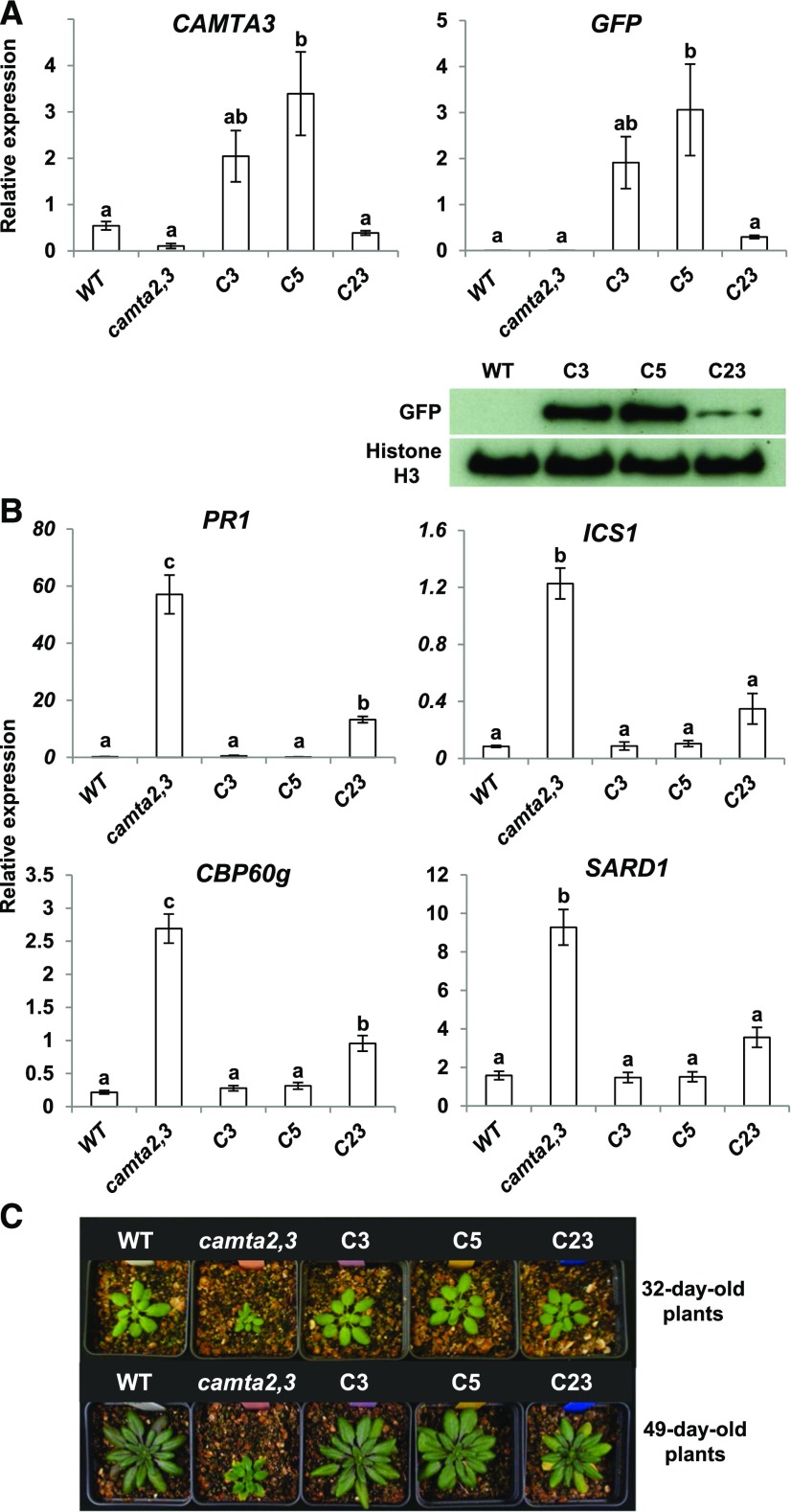
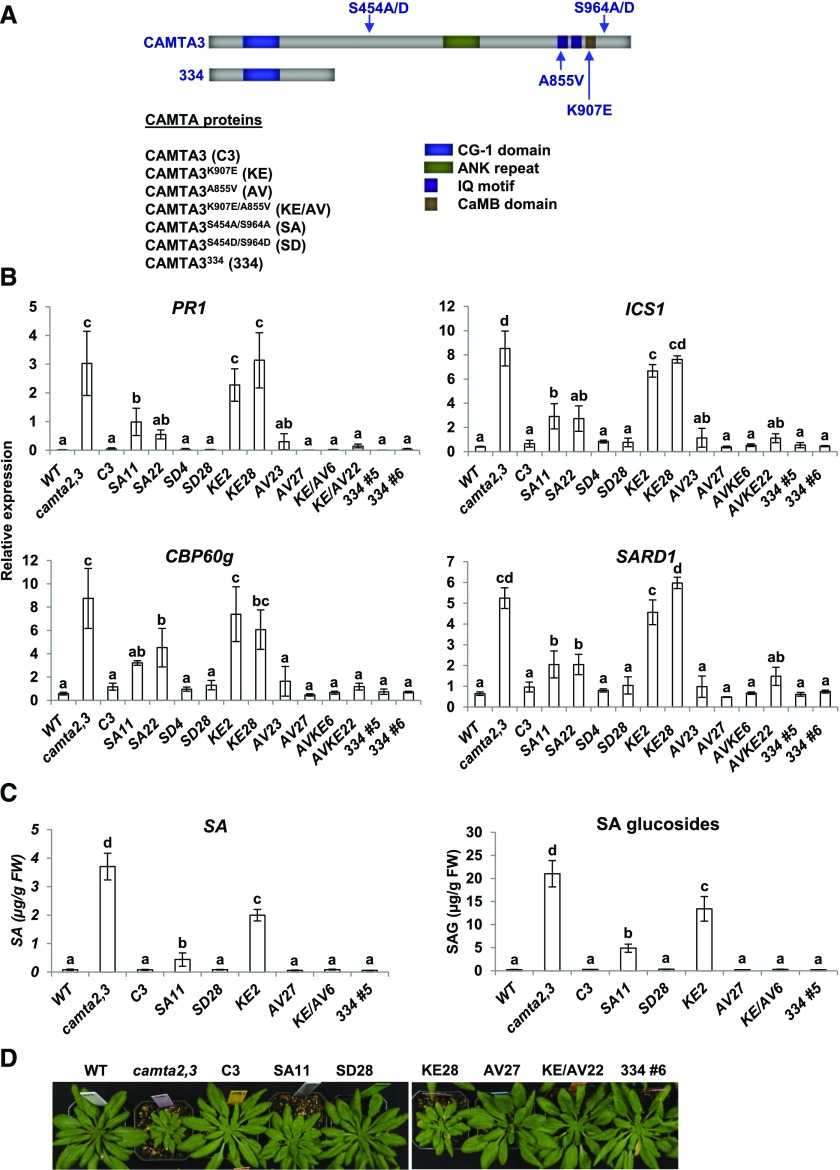
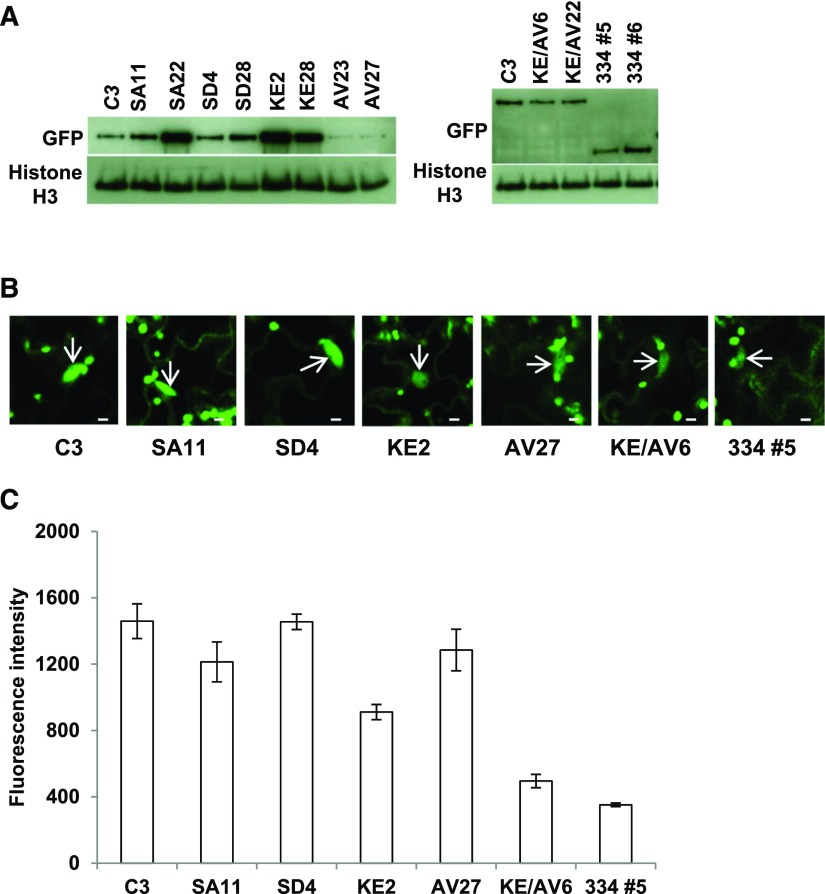
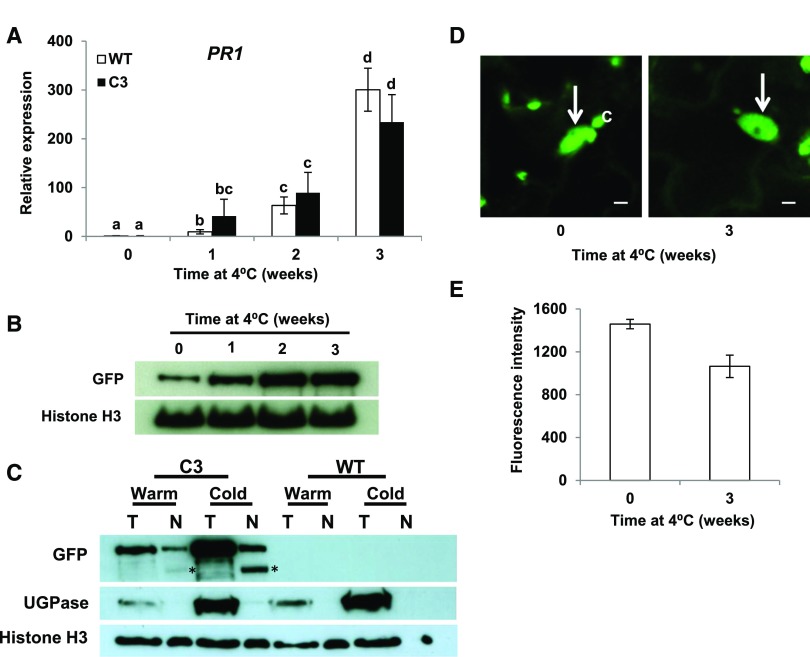
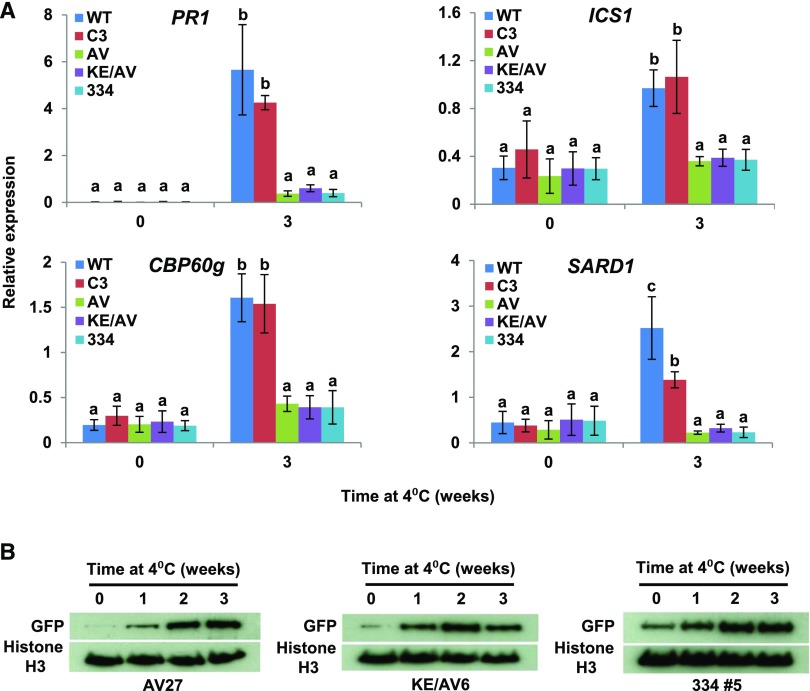
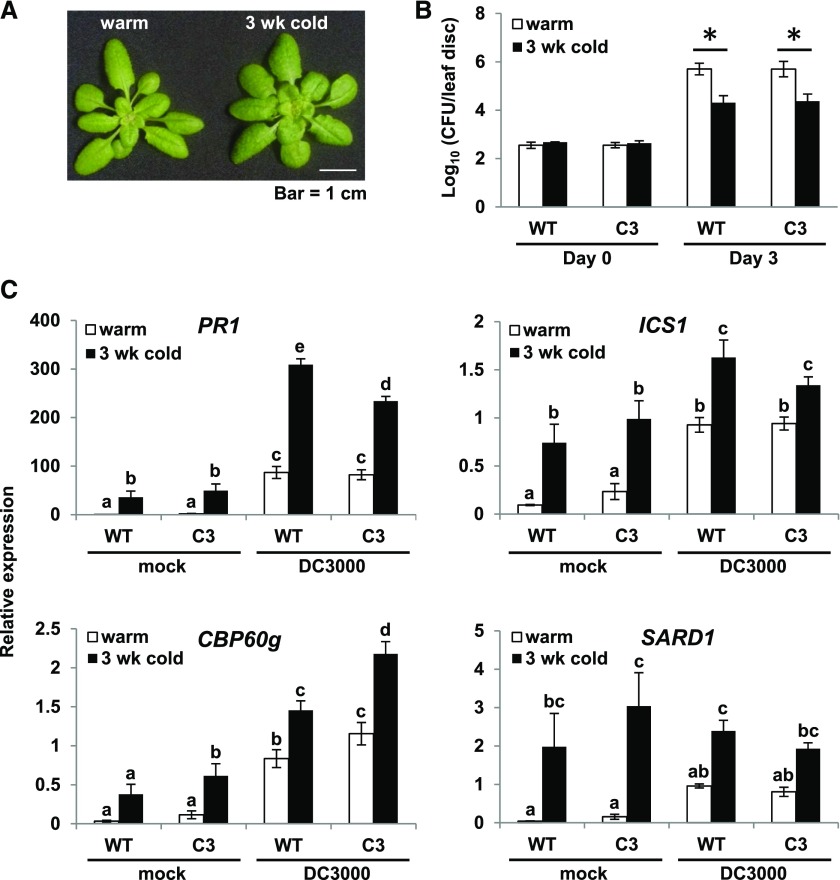
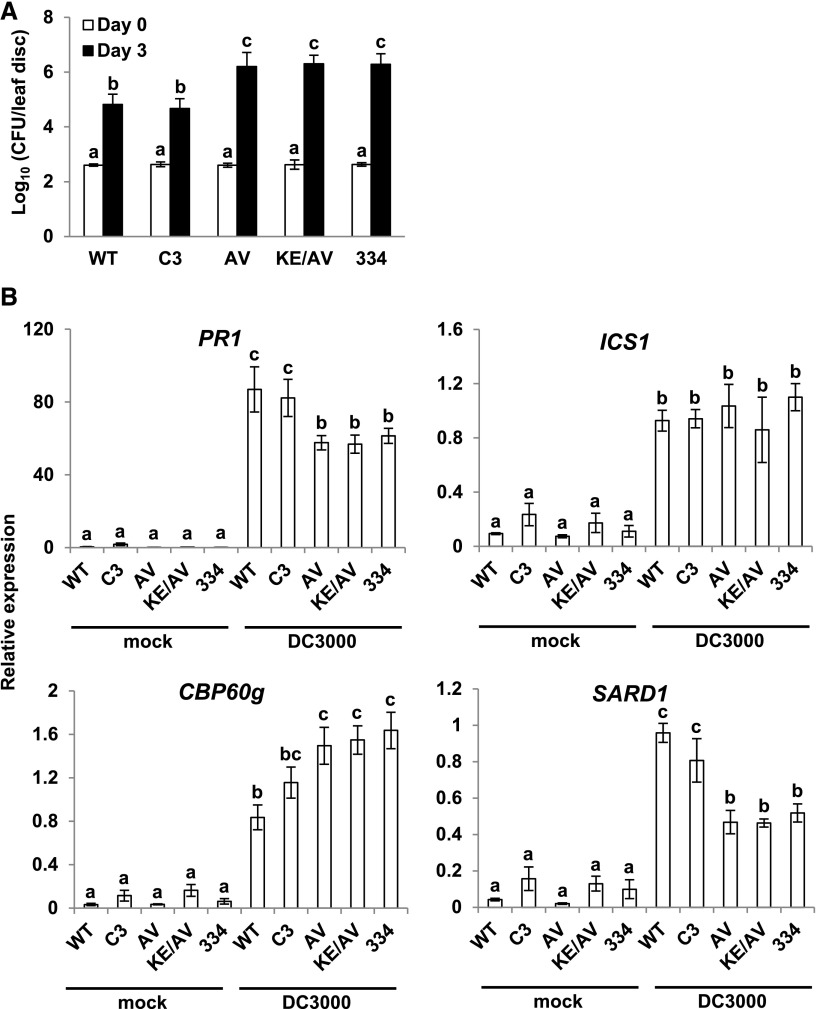
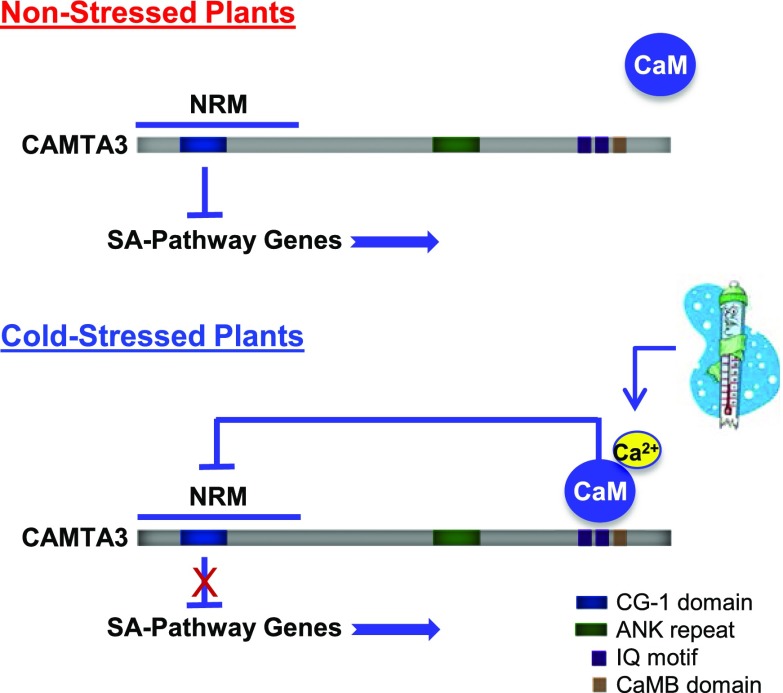
Arabidopsis thaliana calmodulin binding transcription activator (CAMTA) factors repress the expression of genes involved in salicylic acid (SA) biosynthesis and SA-mediated immunity in healthy plants grown at warm temperature (22°C). This repression is overcome in plants exposed to low temperature (4°C) for more than a week and in plants infected by biotrophic and hemibiotrophic pathogens. Here, we present evidence that CAMTA3-mediated repression of SA pathway genes in nonstressed plants involves the action of an N-terminal repression module (NRM) that acts independently of calmodulin (CaM) binding to the IQ and CaM binding (CaMB) domains, a finding that is contrary to current thinking that CAMTA3 repression activity requires binding of CaM to the CaMB domain. Induction of SA pathway genes in response to low temperature did not occur in plants expressing only the CAMTA3-NRM region of the protein. Mutational analysis provided evidence that the repression activity of the NRM was suppressed by action of the IQ and CaMB domains responding to signals generated in response to low temperature. Plants expressing the CAMTA3-NRM region were also impaired in defense against the bacterial hemibiotrophic pathogen Pseudomonas syringae pv tomato DC3000. Our results indicate that the regulation of CAMTA3 repression activity by low temperature and pathogen infection involves related mechanisms, but with distinct differences.
© 2017 American Society of Plant Biologists. All rights reserved.
Figures
References
-
- Bouché N., Scharlat A., Snedden W., Bouchez D., Fromm H. (2002). A novel family of calmodulin-binding transcription activators in multicellular organisms. J. Biol. Chem. 277: 21851–21861. - PubMed
-
- Choi M.S., et al. (2005). Isolation of a calmodulin-binding transcription factor from rice (Oryza sativa L.). J. Biol. Chem. 280: 40820–40831. - PubMed
-
- Clough S.J., Bent A.F. (1998). Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16: 735–743. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
