

Increased Plasmodium falciparum Parasitemia in Non-splenectomized Saimiri sciureus Monkeys Treated with Clodronate Liposomes
- PMID: 28983468
- PMCID: PMC5613086
- DOI: 10.3389/fcimb.2017.00408
Increased Plasmodium falciparum Parasitemia in Non-splenectomized Saimiri sciureus Monkeys Treated with Clodronate Liposomes
Abstract
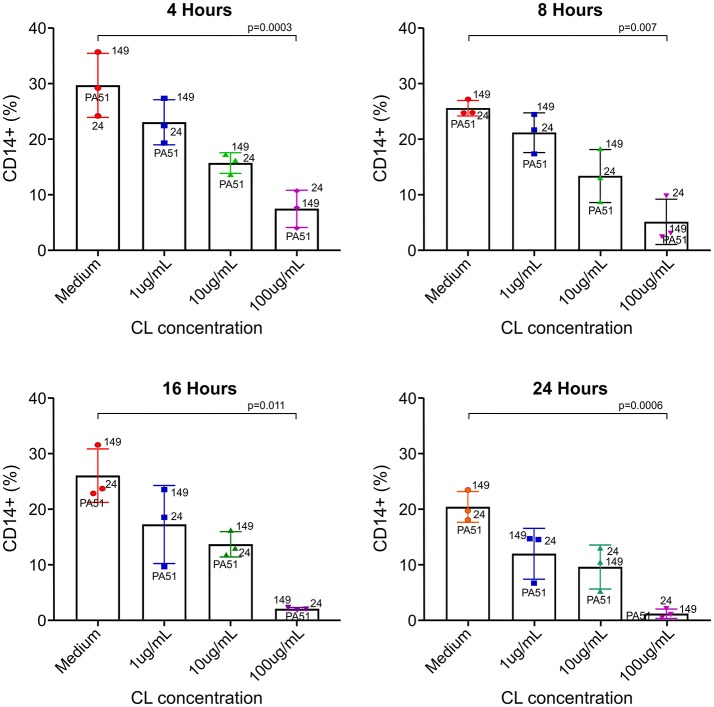
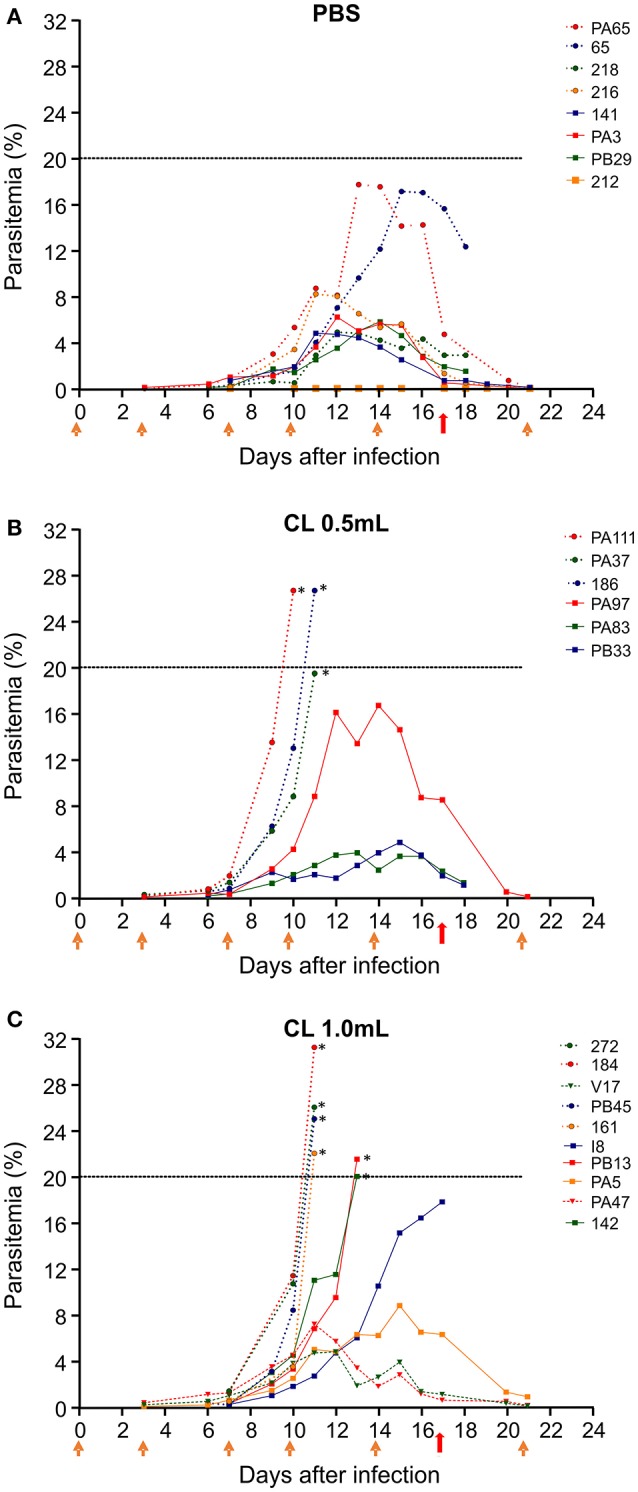
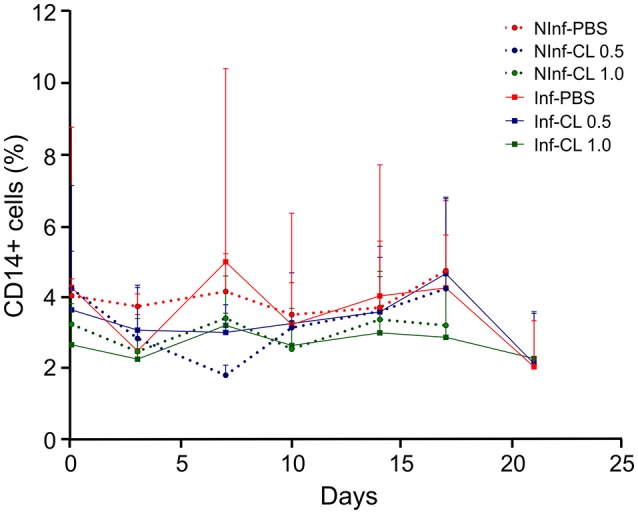
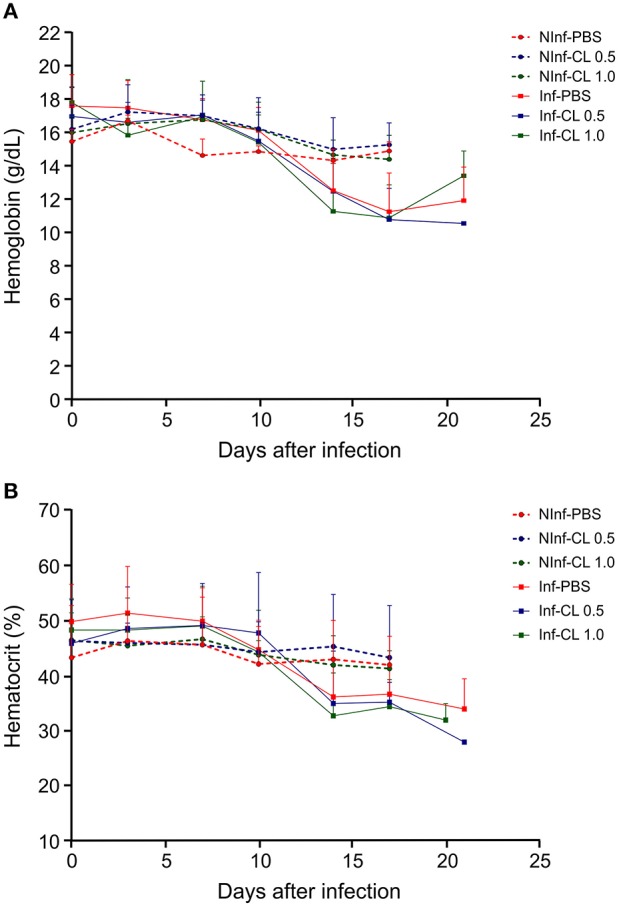
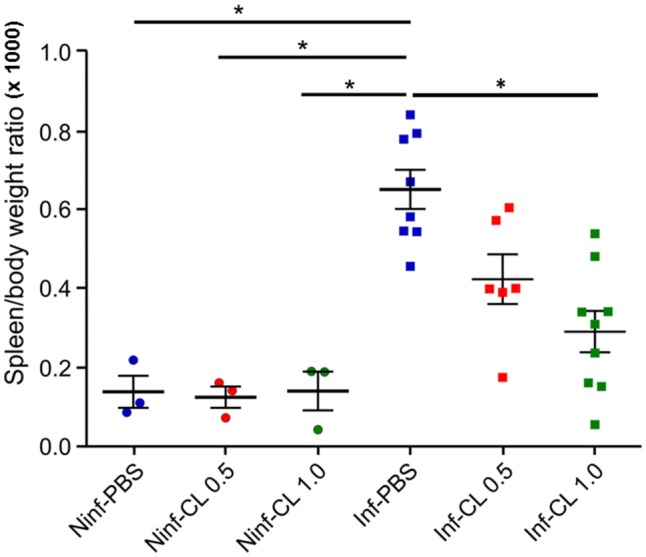
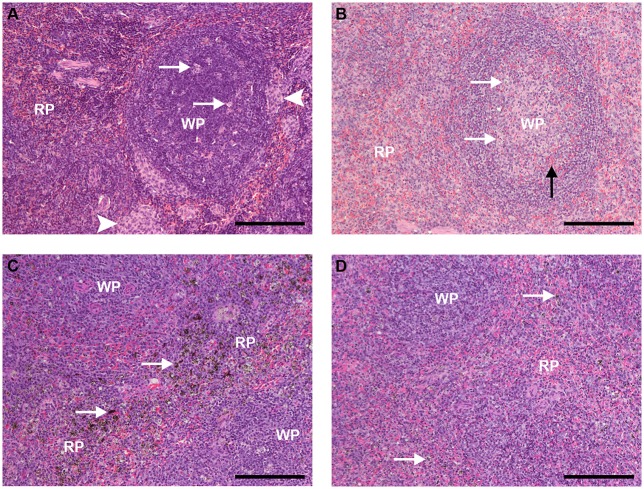
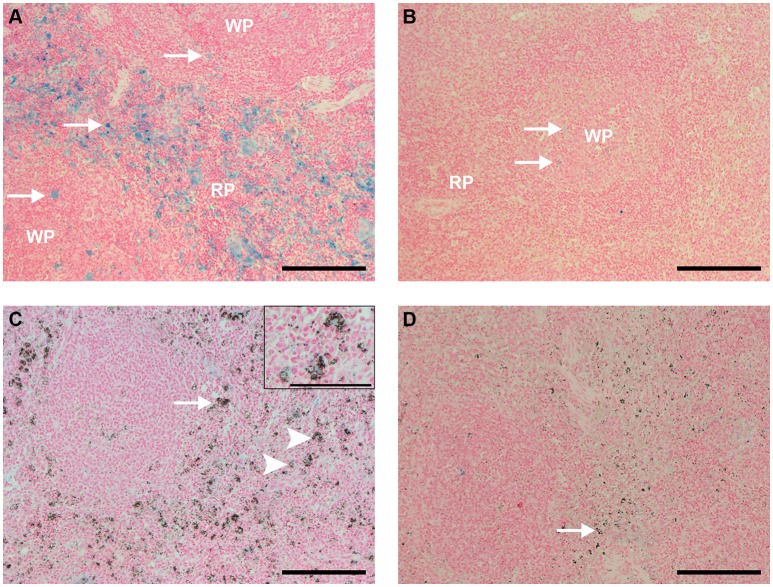
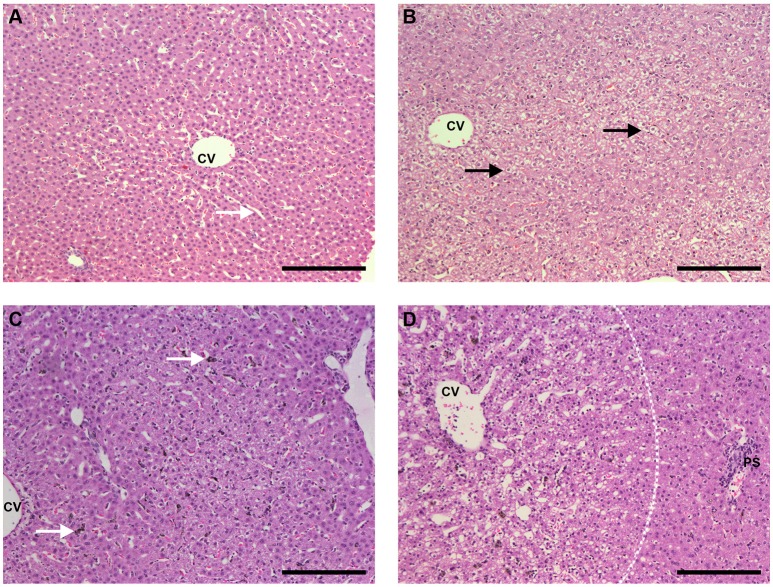
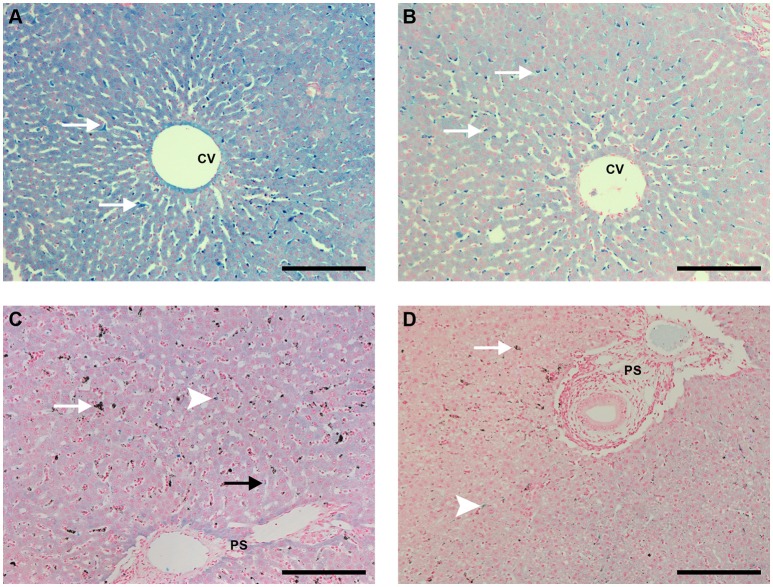
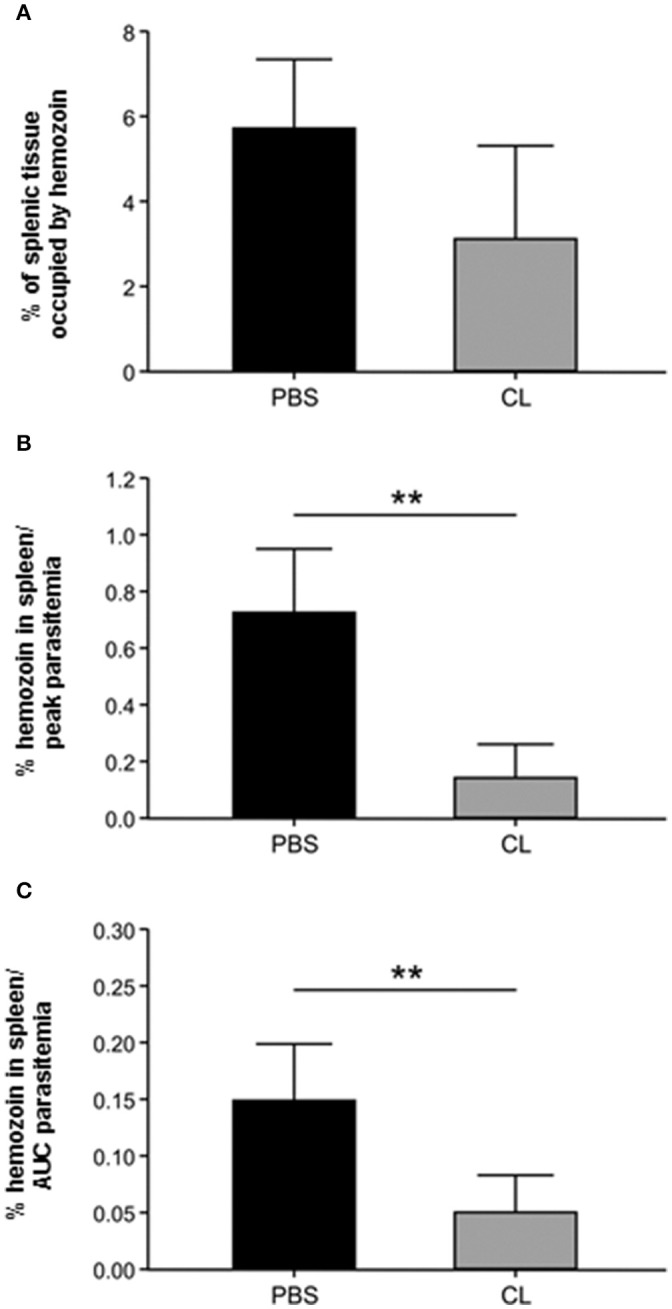
A major constraint in the study of Plasmodium falciparum malaria, including vaccine development, lies on the parasite's strict human host specificity and therefore the shortage of animal experimental models able to harbor human plasmodia. The best experimental models are neo-tropical primates of the genus Saimiri and Aotus, but they require splenectomy to reduce innate defenses for achieving high and consistent parasitemias, an important limitation. Clodronate-liposomes (CL) have been successfully used to deplete monocytes/macrophages in several experimental models. We investigated whether a reduction in the numbers of phagocytic cells by CL would improve the development of P. falciparum parasitemia in non-splenectomized Saimiri sciureus monkeys. Depletion of S. sciureus splenocytes after in vitro incubation with CL was quantified using anti-CD14 antibodies and flow cytometry. Non-infected and P. falciparum-infected S. sciureus were injected intravenously twice a week with either CL at either 0.5 or 1 mL (5 mg/mL) or phosphate buffered saline (PBS). Animals were monitored during infection and treated with mefloquine. After treatment and euthanasia, spleen and liver were collected for histological analysis. In vitro CL depleted S. sciureus splenic monocyte/macrophage population in a dose- and time-dependent manner. In vivo, half of P. falciparum-infected S. sciureus treated with CL 0.5 mL, and two-thirds of those treated with CL 1 mL developed high parasitemias requiring mefloquine treatment, whereas all control animals were able to self-control parasitemia without the need for antimalarial treatment. CL-treated infected S. sciureus showed a marked decrease in the degree of splenomegaly despite higher parasitemias, compared to PBS-treated animals. Histological evidence of partial monocyte/macrophage depletion, decreased hemozoin phagocytosis and decreased iron recycling was observed in both the spleen and liver of CL-treated infected S. sciureus. CL is capable of promoting higher parasitemia in P. falciparum-infected S. sciureus, associated with evidence of partial macrophage depletion in the spleen and liver. Macrophage depletion by CL is therefore a practical and viable alternative to surgical splenectomy in this experimental model.
Keywords: Plasmodium falciparum; Saimiri sciureus; clodronate liposomes; liver; macrophages; malaria; spleen.
Figures
References
-
- Abbas A. K., Lichtman A. H., Pillai S. (2008). Imunologia Celular e Molecular, 6th Edn. Rio de Janeiro: Elsevier.
-
- Alves F. A., Pelajo-Machado M., Totino P. R., Souza M. T., Gonçalves E. C., Schneider M. P., et al. (2015). Splenic architecture disruption and parasite-induced splenocyte activation and anergy in Plasmodium falciparum-infected Saimiri sciureus monkeys. Malar. J. 14:128. 10.1186/s12936-015-0641-3 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials