

Engineering the Genetic Code in Cells and Animals: Biological Considerations and Impacts
- PMID: 28984438
- PMCID: PMC5698093
- DOI: 10.1021/acs.accounts.7b00376
Engineering the Genetic Code in Cells and Animals: Biological Considerations and Impacts
Abstract
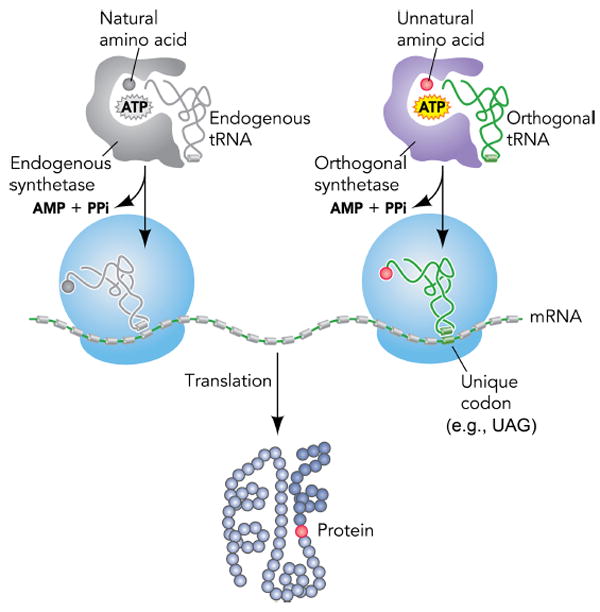
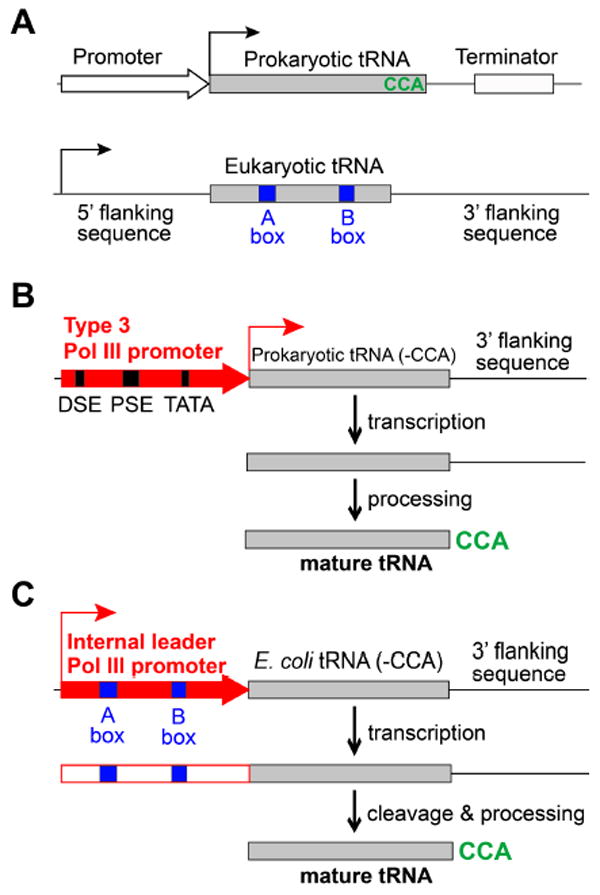
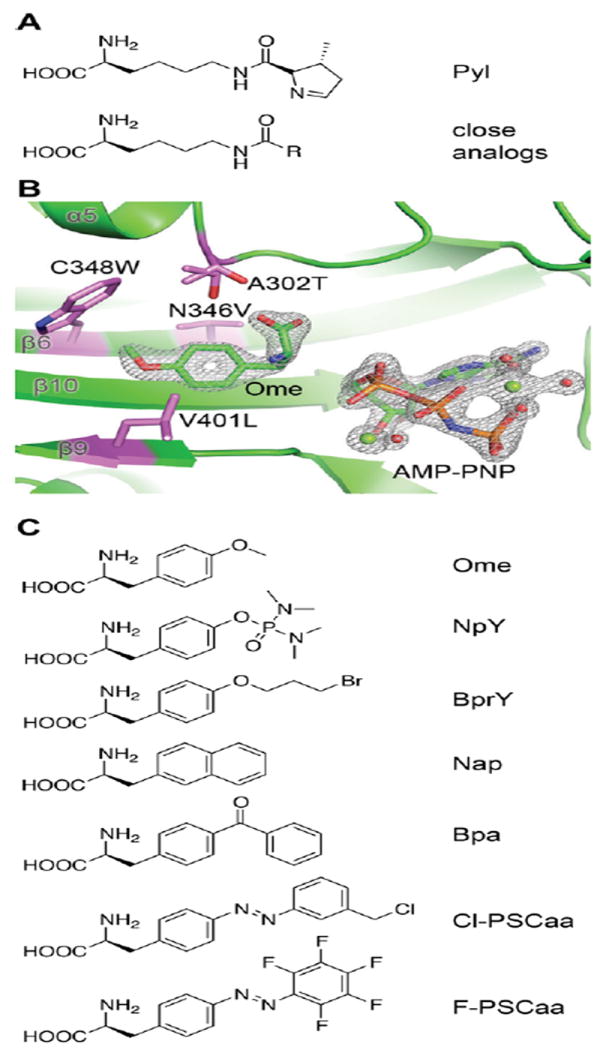
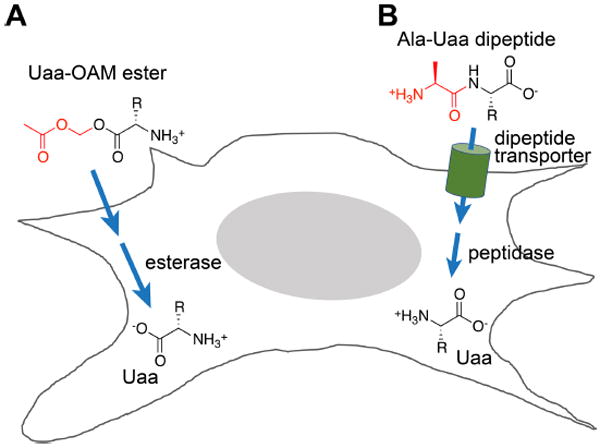
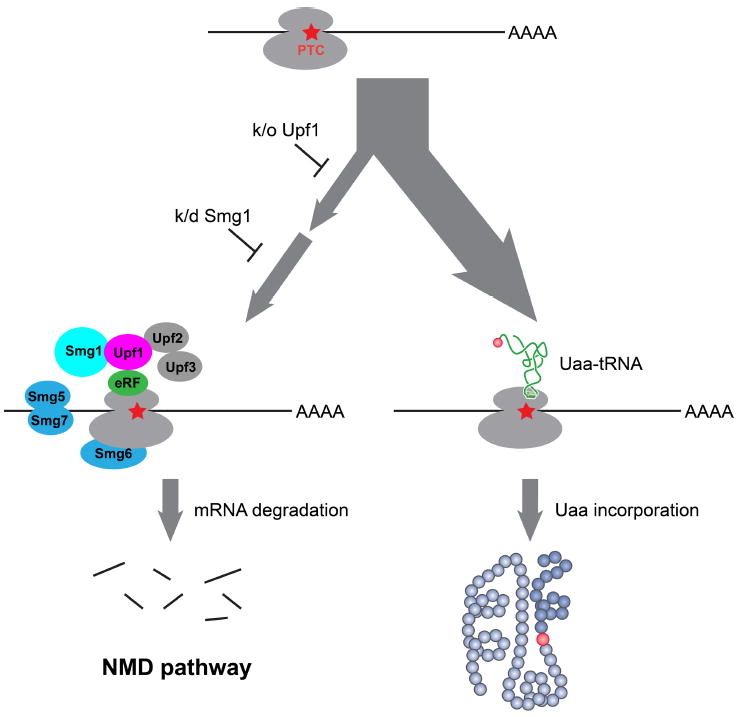
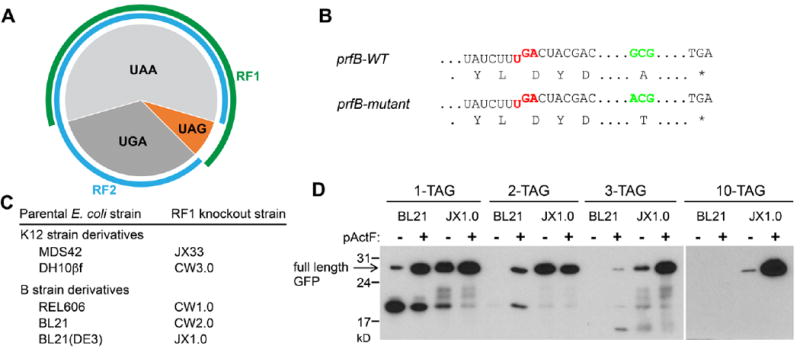
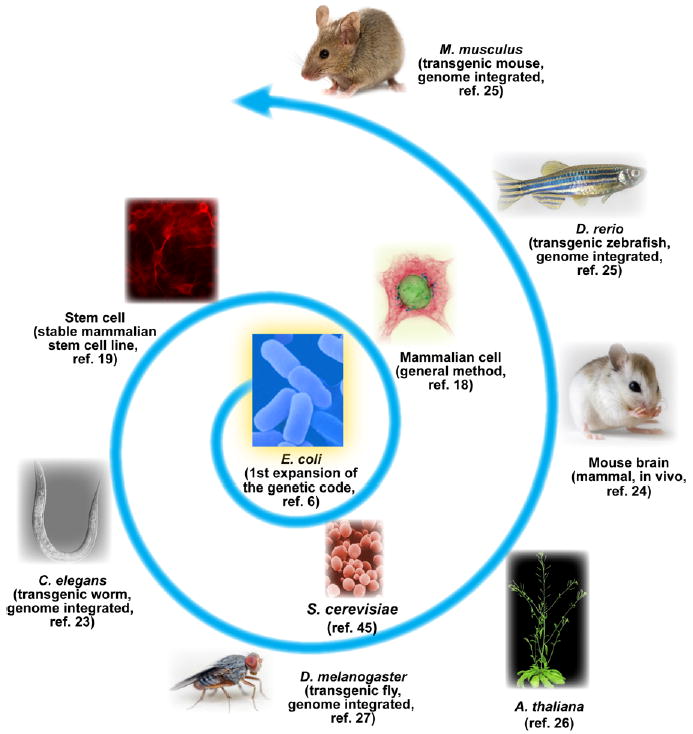
Expansion of the genetic code allows unnatural amino acids (Uaas) to be site-specifically incorporated into proteins in live biological systems, thus enabling novel properties selectively introduced into target proteins in vivo for basic biological studies and for engineering of novel biological functions. Orthogonal components including tRNA and aminoacyl-tRNA synthetase (aaRS) are expressed in live cells to decode a unique codon (often the amber stop codon UAG) as the desired Uaa. Initially developed in E. coli, this methodology has now been expanded in multiple eukaryotic cells and animals. In this Account, we focus on addressing various biological challenges for rewriting the genetic code, describing impacts of code expansion on cell physiology and discussing implications for fundamental studies of code evolution. Specifically, a general method using the type-3 polymerase III promoter was developed to efficiently express prokaryotic tRNAs as orthogonal tRNAs and a transfer strategy was devised to generate Uaa-specific aaRS for use in eukaryotic cells and animals. The aaRSs have been found to be highly amenable for engineering substrate specificity toward Uaas that are structurally far deviating from the native amino acid, dramatically increasing the stereochemical diversity of Uaas accessible. Preparation of the Uaa in ester or dipeptide format markedly increases the bioavailability of Uaas to cells and animals. Nonsense-mediated mRNA decay (NMD), an mRNA surveillance mechanism of eukaryotic cells, degrades mRNA containing a premature stop codon. Inhibition of NMD increases Uaa incorporation efficiency in yeast and Caenorhabditis elegans. In bacteria, release factor one (RF1) competes with the orthogonal tRNA for the amber stop codon to terminate protein translation, leading to low Uaa incorporation efficiency. Contradictory to the paradigm that RF1 is essential, it is discovered that RF1 is actually nonessential in E. coli. Knockout of RF1 dramatically increases Uaa incorporation efficiency and enables Uaa incorporation at multiple sites, making it feasible to use Uaa for directed evolution. Using these strategies, the genetic code has been effectively expanded in yeast, mammalian cells, stem cells, worms, fruit flies, zebrafish, and mice. It is also intriguing to find out that the legitimate UAG codons terminating endogenous genes are not efficiently suppressed by the orthogonal tRNA/aaRS in E. coli. Moreover, E. coli responds to amber suppression pressure promptly using transposon insertion to inactivate the introduced orthogonal aaRS. Persistent amber suppression evading transposon inactivation leads to global proteomic changes with a notable up-regulation of a previously uncharacterized protein YdiI, for which an unexpected function of expelling plasmids is discovered. Genome integration of the orthogonal tRNA/aaRS in mice results in minor changes in RNA transcripts but no significant physiological impairment. Lastly, the RF1 knockout E. coli strains afford a previously unavailable model organism for studying otherwise intractable questions on code evolution in real time in the laboratory. We expect that genetically encoding Uaas in live systems will continue to unfold new questions and directions for studying biology in vivo, investigating the code itself, and reprograming genomes for synthetic biology.
Figures
References
-
- Wang L, Schultz PG. Expanding the genetic code. Chem Commun. 2002:1–11. - PubMed
-
- Wang L, Schultz PG. Expanding the genetic code. Angew Chem Int Ed Engl. 2005;44:34–66. - PubMed
-
- Wang L, Magliery TJ, Liu DR, Schultz PG. A new functional suppressor tRNA/aminoacyl-tRNA synthetase pair for the in vivo incorporation of unnatural amino acids into proteins. J Am Chem Soc. 2000;122:5010–5011.
-
- Wang L, Schultz PG. A general approach for the generation of orthogonal tRNAs. Chem Biol. 2001;8:883–890. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
