

B cell tolerance to epidermal ribonuclear-associated neo-autoantigen in vivo
- PMID: 28984923
- PMCID: PMC5758376
- DOI: 10.1111/cei.13066
B cell tolerance to epidermal ribonuclear-associated neo-autoantigen in vivo
Abstract
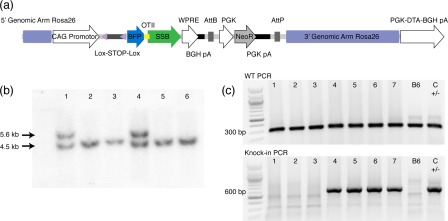
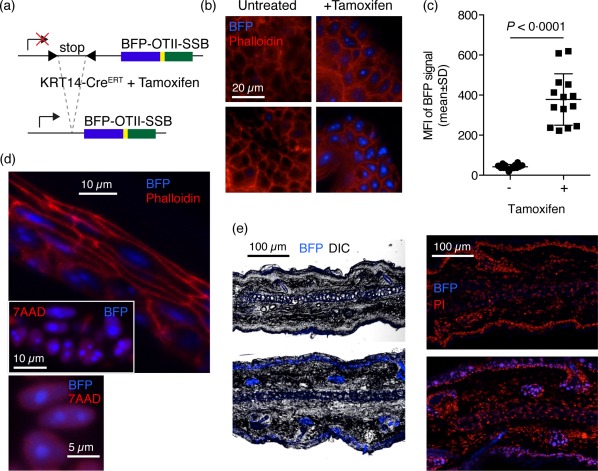
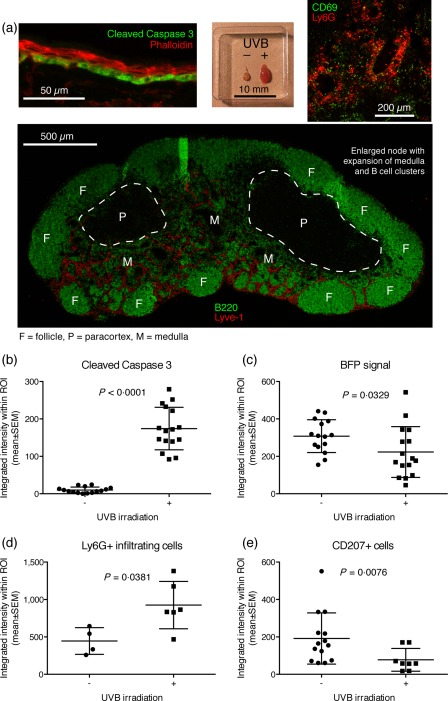
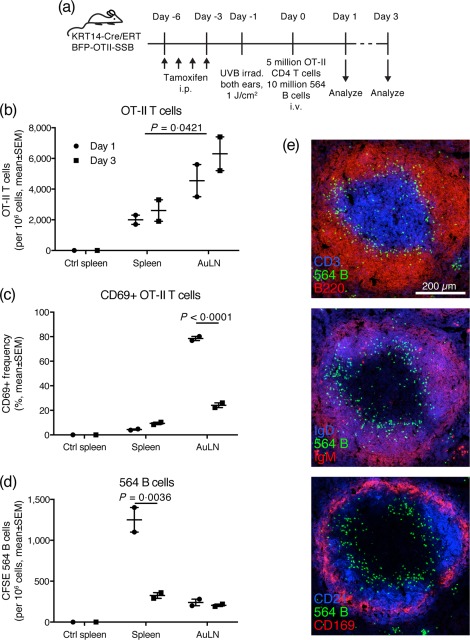
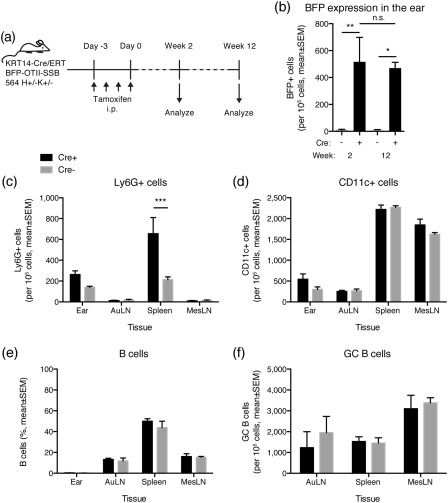
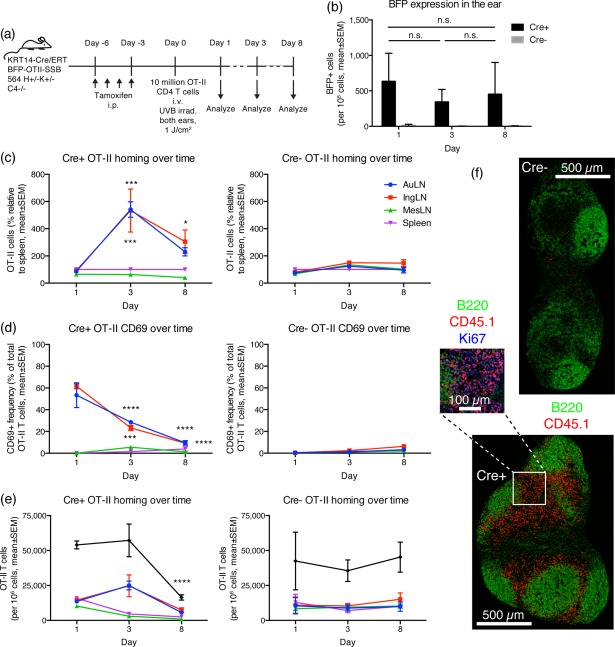
Defining how self-antigens are perceived by the immune system is pivotal to understand how tolerance is maintained under homeostatic conditions. Clinically relevant, natural autoantigens targeted by autoantibodies, in e.g. systemic lupus erythematosus (SLE), commonly have an intrinsic ability to engage not only the B cell receptor (BCR), but also a co-stimulatory pathway in B cells, such as the Toll-like receptor (TLR)-7 pathway. Here we developed a novel mouse model displaying inducible expression of a fluorescent epidermal neo-autoantigen carrying an OT-II T cell epitope, B cell antigen and associated ribonucleic acids capable of stimulating TLR-7. The neo-autoantigen was expressed in skin, but did not drain in intact form into draining lymph nodes, even after ultraviolet B (UVB)-stimulated induction of apoptosis in the basal layer. Adoptively transferred autoreactive B cells were excluded follicularly and perished at the T-B border in the spleen, preventing their recirculation and encounter with antigen peripherally. This transitional check-point was bypassed by crossing the reporter to a BCR knock-in line on a C4-deficient background. Adoptively transferred OT-II T cells homed rapidly into cutaneous lymph nodes and up-regulated CD69. Surprisingly, however, tolerance was not broken, as the T cells subsequently down-regulated activation markers and contracted. Our results highlight how sequestration of intracellular and peripheral antigen, the transitional B cell tolerance check-point and T cell regulation co-operate to maintain immunological tolerance in vivo.
Keywords: B cell; autoimmunity; skin antigen; systemic lupus erythematosus; tolerance.
© 2017 British Society for Immunology.
Figures
Similar articles
-
B and T cell tolerance and autoimmunity in autoantibody transgenic mice.Int Immunol. 2002 Aug;14(8):963-71. doi: 10.1093/intimm/dxf064. Int Immunol. 2002. PMID: 12147633
-
A major B cell epitope present on the apoptotic but not the intact form of the U1-70-kDa ribonucleoprotein autoantigen.J Immunol. 2004 Jan 1;172(1):709-16. doi: 10.4049/jimmunol.172.1.709. J Immunol. 2004. PMID: 14688384
-
Cutting edge: CD25+ regulatory T cells prevent expansion and induce apoptosis of B cells specific for tissue autoantigens.J Immunol. 2008 Oct 1;181(7):4447-51. doi: 10.4049/jimmunol.181.7.4447. J Immunol. 2008. PMID: 18802046
-
Molecular mimicry and the role of B lymphocytes in the processing of autoantigens.Cell Mol Life Sci. 2000 Apr;57(4):561-8. doi: 10.1007/PL00000718. Cell Mol Life Sci. 2000. PMID: 11130456 Free PMC article. Review.
-
Dysregulated Lymphoid Cell Populations in Mouse Models of Systemic Lupus Erythematosus.Clin Rev Allergy Immunol. 2017 Oct;53(2):181-197. doi: 10.1007/s12016-017-8605-8. Clin Rev Allergy Immunol. 2017. PMID: 28500565 Review.
Cited by
-
Mechanosensing by Peyer's patch stroma regulates lymphocyte migration and mucosal antibody responses.Nat Immunol. 2019 Nov;20(11):1506-1516. doi: 10.1038/s41590-019-0505-z. Epub 2019 Oct 14. Nat Immunol. 2019. PMID: 31611698 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
