

Reactive metabolites of acetaminophen activate and sensitize the capsaicin receptor TRPV1
- PMID: 28986540
- PMCID: PMC5630573
- DOI: 10.1038/s41598-017-13054-3
Reactive metabolites of acetaminophen activate and sensitize the capsaicin receptor TRPV1
Abstract
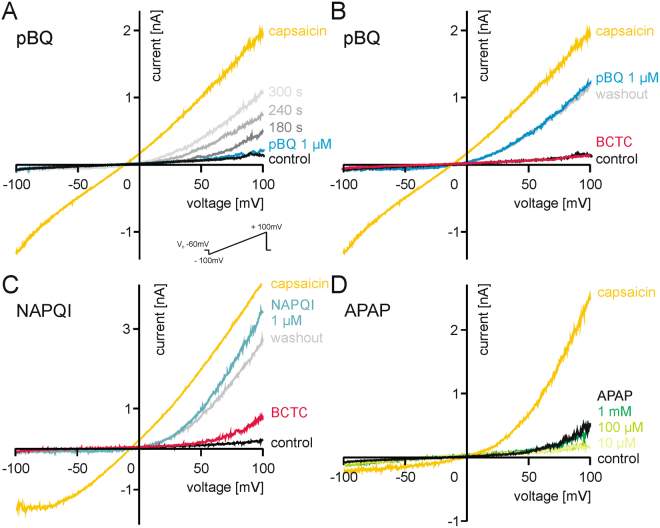
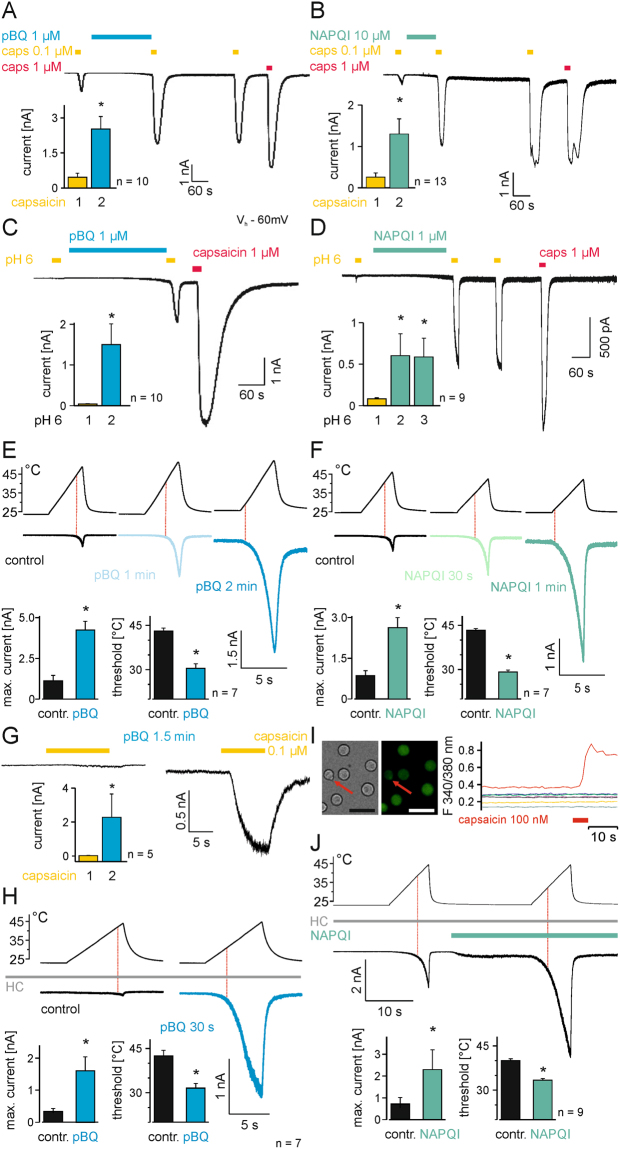
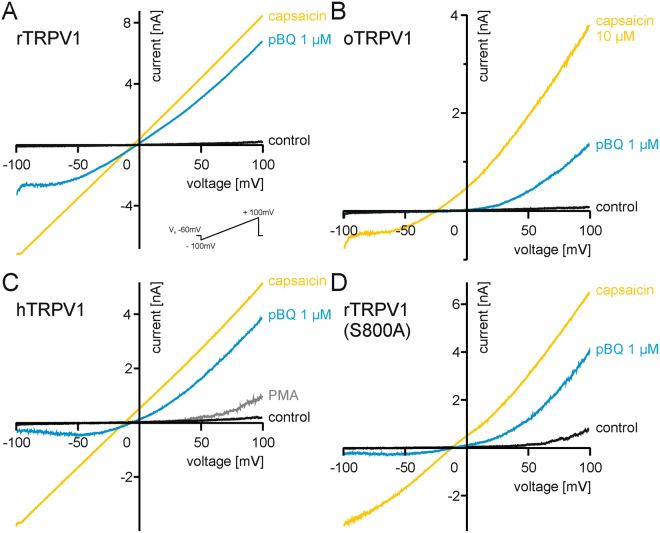
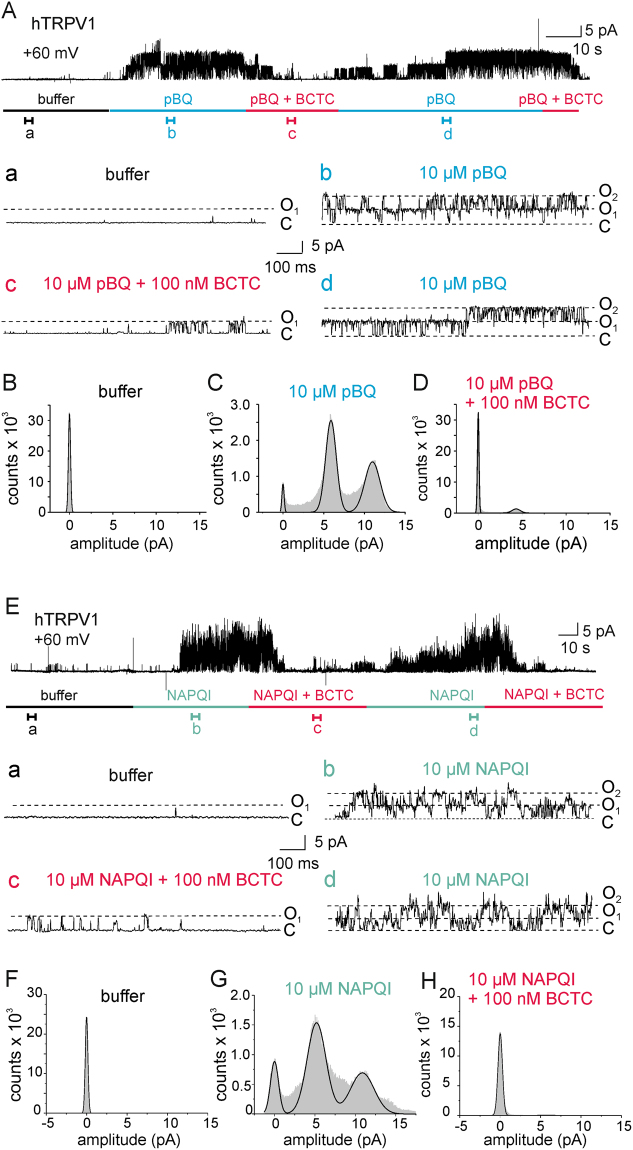
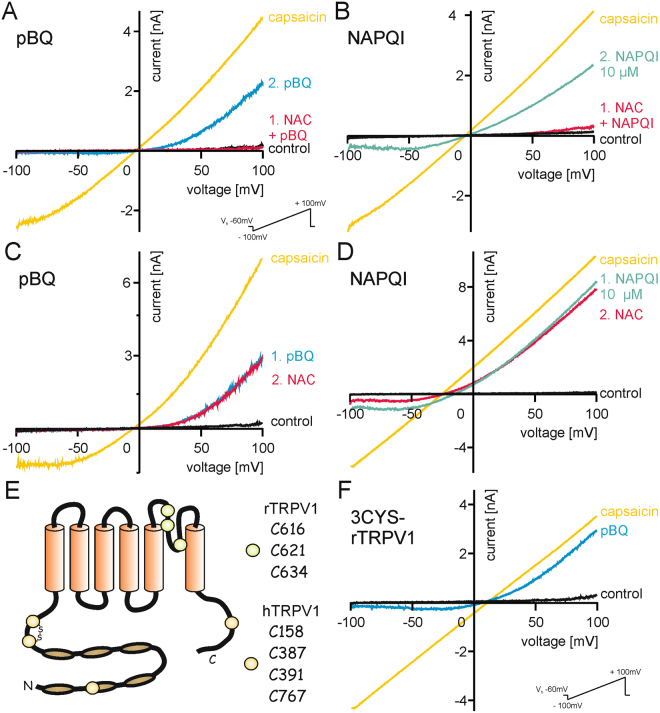
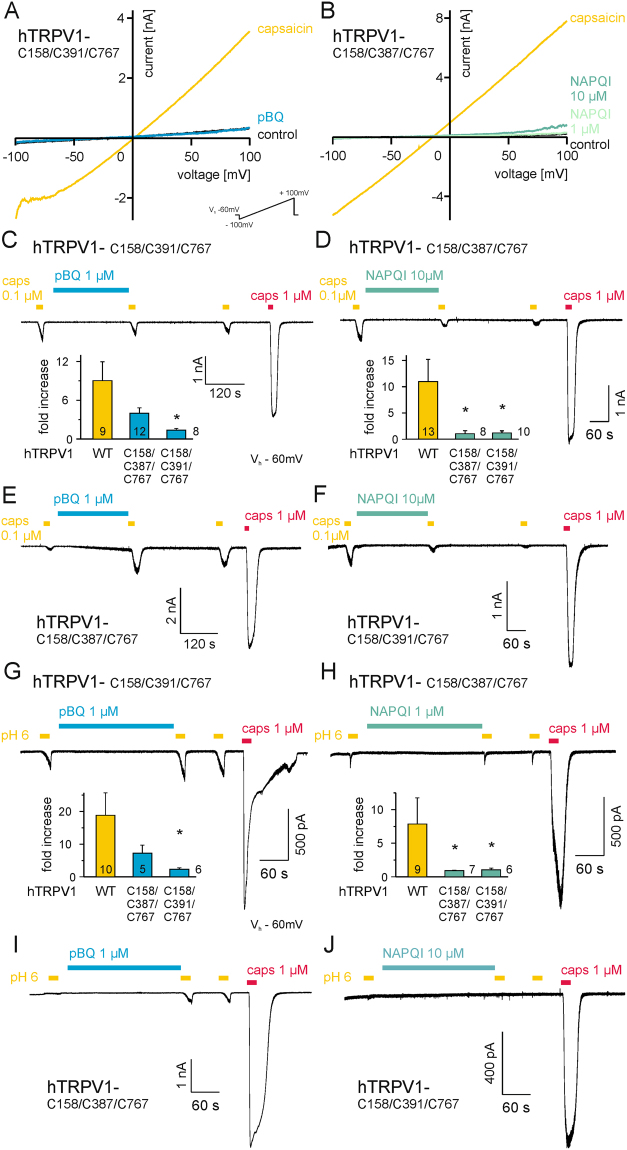
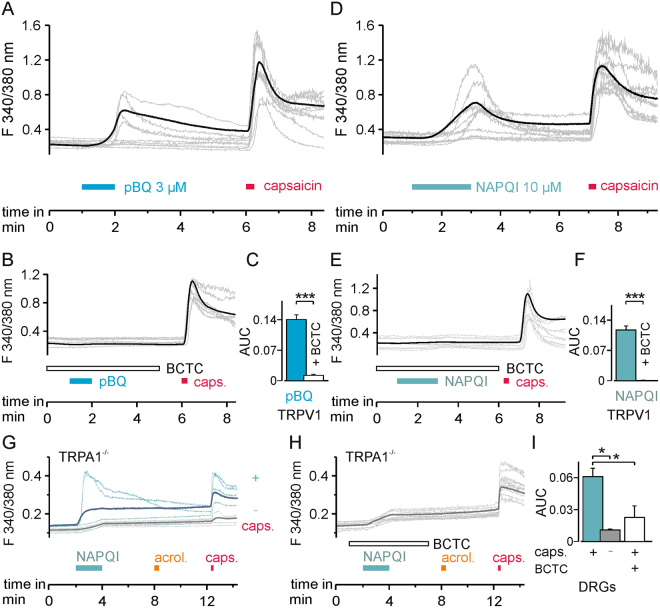
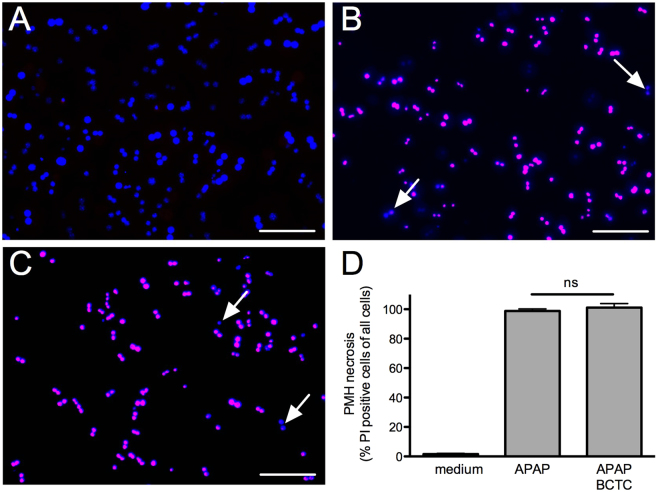
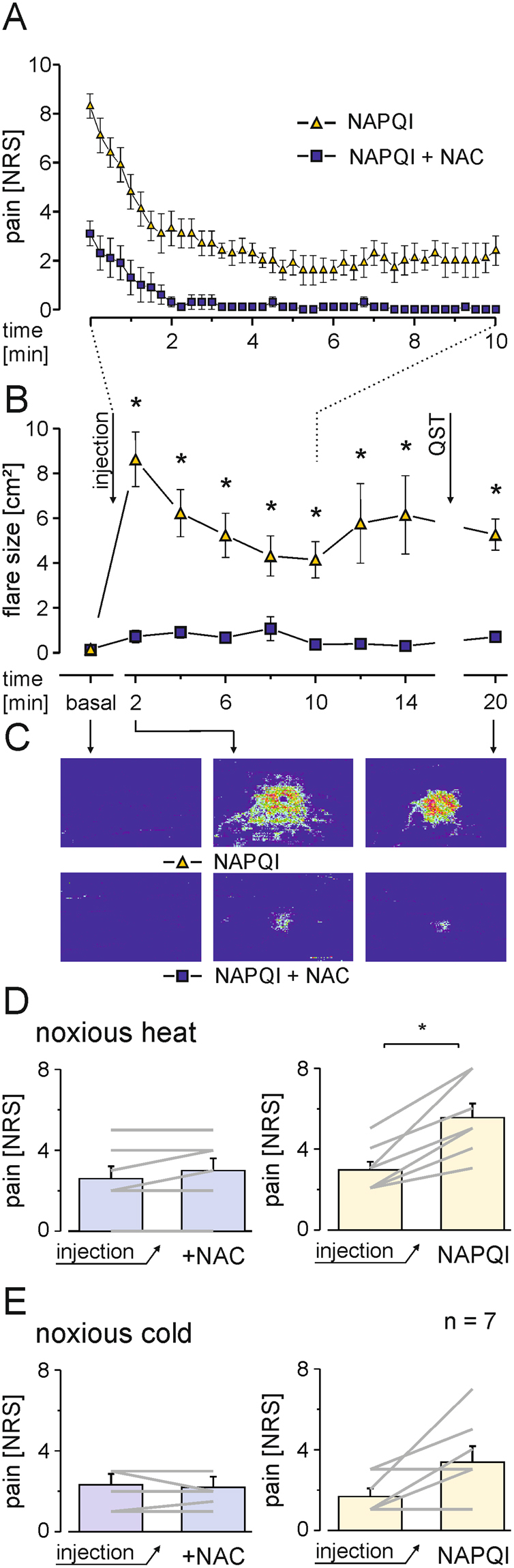
The irritant receptor TRPA1 was suggested to mediate analgesic, antipyretic but also pro-inflammatory effects of the non-opioid analgesic acetaminophen, presumably due to channel activation by the reactive metabolites parabenzoquinone (pBQ) and N-acetyl-parabenzoquinonimine (NAPQI). Here we explored the effects of these metabolites on the capsaicin receptor TRPV1, another redox-sensitive ion channel expressed in sensory neurons. Both pBQ and NAPQI, but not acetaminophen irreversibly activated and sensitized recombinant human and rodent TRPV1 channels expressed in HEK 293 cells. The reducing agents dithiothreitol and N-acetylcysteine abolished these effects when co-applied with the metabolites, and both pBQ and NAPQI failed to gate TRPV1 following substitution of the intracellular cysteines 158, 391 and 767. NAPQI evoked a TRPV1-dependent increase in intracellular calcium and a potentiation of heat-evoked currents in mouse spinal sensory neurons. Although TRPV1 is expressed in mouse hepatocytes, inhibition of TRPV1 did not alleviate acetaminophen-induced hepatotoxicity. Finally, intracutaneously applied NAPQI evoked burning pain and neurogenic inflammation in human volunteers. Our data demonstrate that pBQ and NAQPI activate and sensitize TRPV1 by interacting with intracellular cysteines. While TRPV1 does not seem to mediate acetaminophen-induced hepatotoxicity, our data identify TRPV1 as a target of acetaminophen with a potential relevance for acetaminophen-induced analgesia, antipyresia and inflammation.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Mitchell JR, Jollow DJ, Potter WZ, Gillette JR, Brodie BB. Acetaminophen-induced hepatic necrosis. IV. Protective role of glutathione. The Journal of pharmacology and experimental therapeutics. 1973;187:211–217. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
