

NMDA Receptors in the Central Nervous System
- PMID: 28986865
- PMCID: PMC7325486
- DOI: 10.1007/978-1-4939-7321-7_1
NMDA Receptors in the Central Nervous System
Abstract
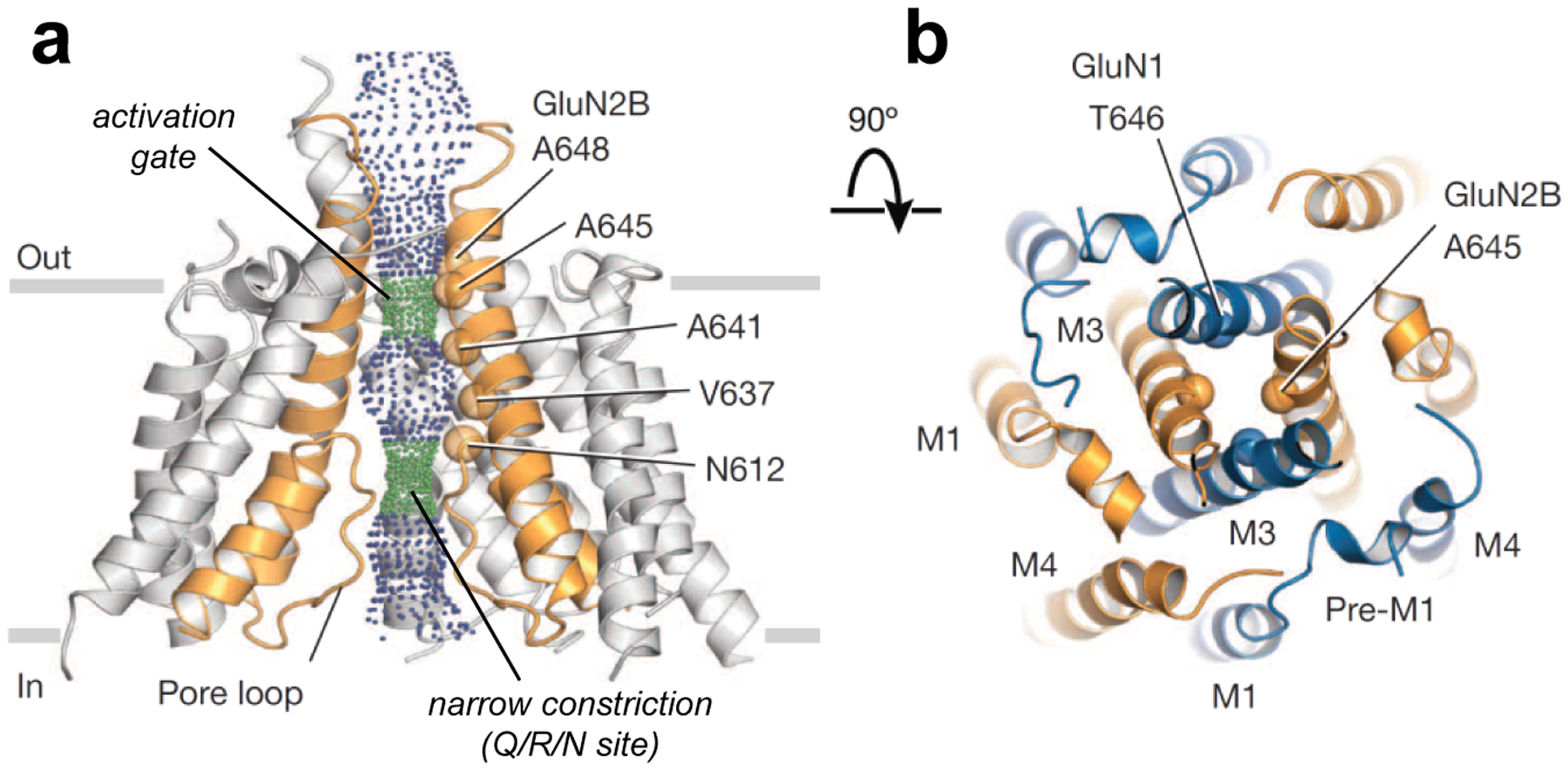
NMDA-type glutamate receptors are ligand-gated ion channels that mediate a major component of excitatory neurotransmission in the central nervous system (CNS). They are widely distributed at all stages of development and are critically involved in normal brain functions, including neuronal development and synaptic plasticity. NMDA receptors are also implicated in the pathophysiology of numerous neurological and psychiatric disorders, such as ischemic stroke, traumatic brain injury, Alzheimer's disease, epilepsy, mood disorders, and schizophrenia. For these reasons, NMDA receptors have been intensively studied in the past several decades to elucidate their physiological roles and to advance them as therapeutic targets. Seven NMDA receptor subunits exist that assemble into a diverse array of tetrameric receptor complexes, which are differently regulated, have distinct regional and developmental expression, and possess a wide range of functional and pharmacological properties. The diversity in subunit composition creates NMDA receptor subtypes with distinct physiological roles across neuronal cell types and brain regions, and enables precise tuning of synaptic transmission. Here, we will review the relationship between NMDA receptor structure and function, the diversity and significance of NMDA receptor subtypes in the CNS, as well as principles and rules by which NMDA receptors operate in the CNS under normal and pathological conditions.
Keywords: Disease; Ion channel; Ionotropic glutamate receptor; NMDA; Neurotransmitter; Regulation; Structure-function; Synaptic transmission.
Figures
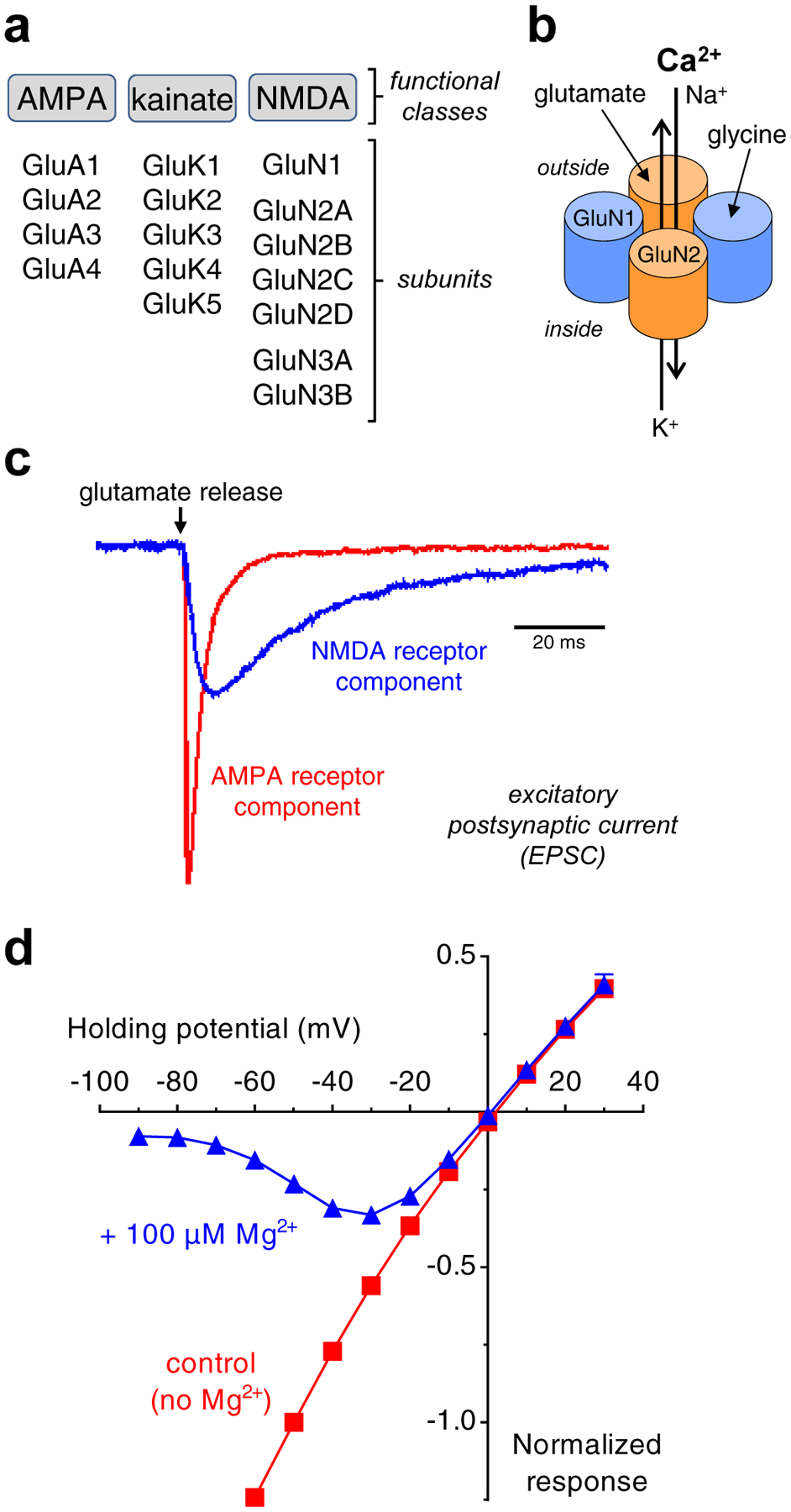
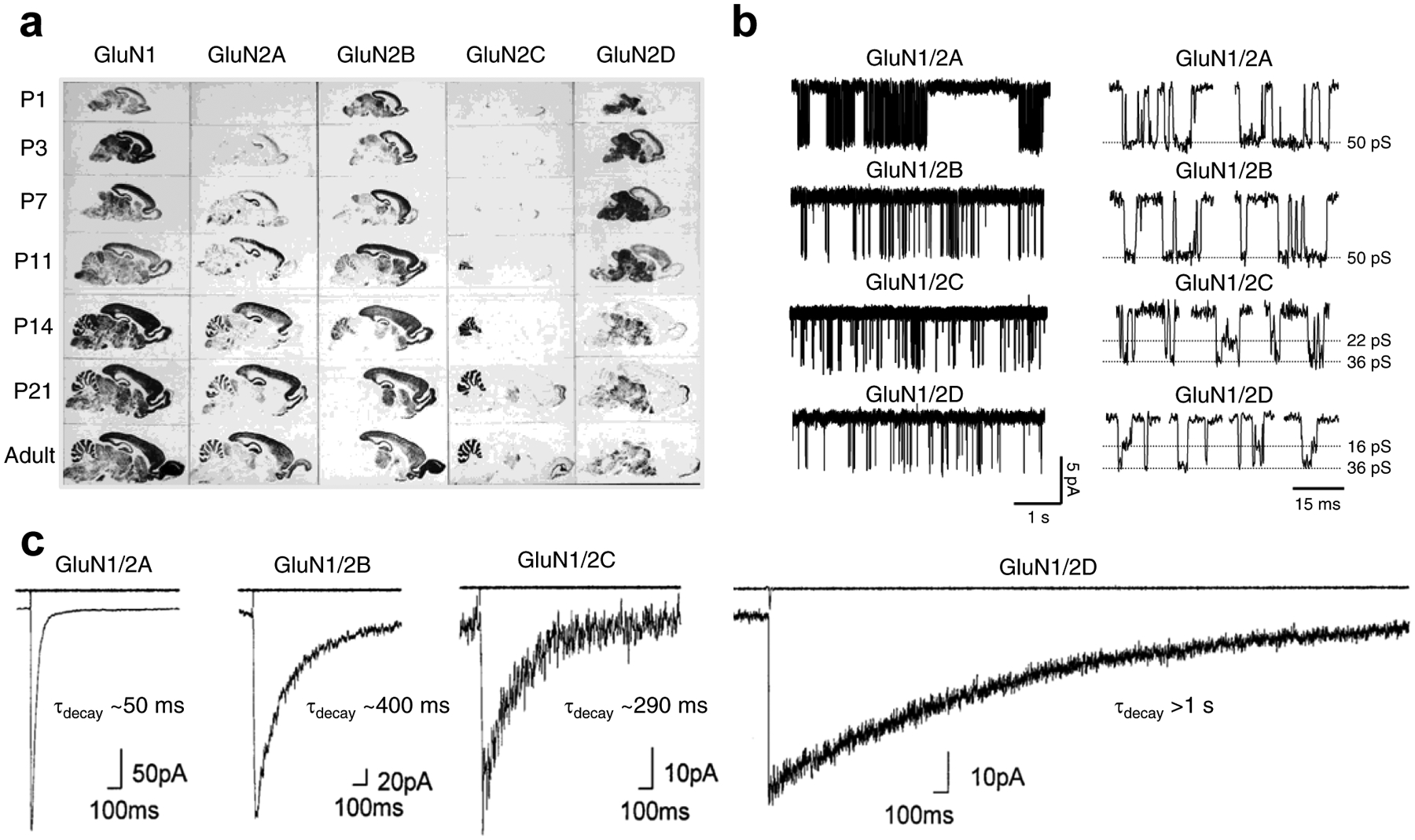
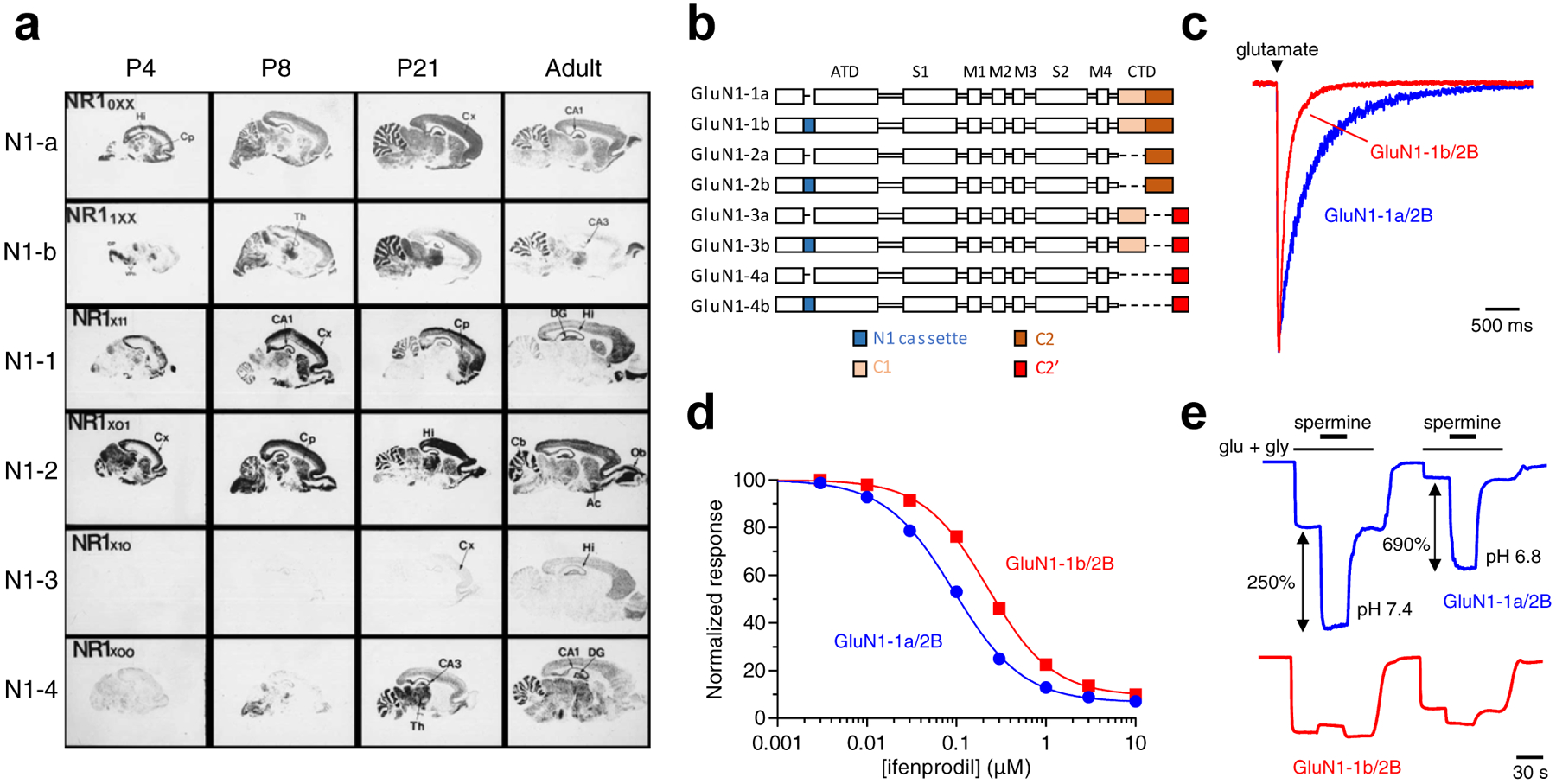

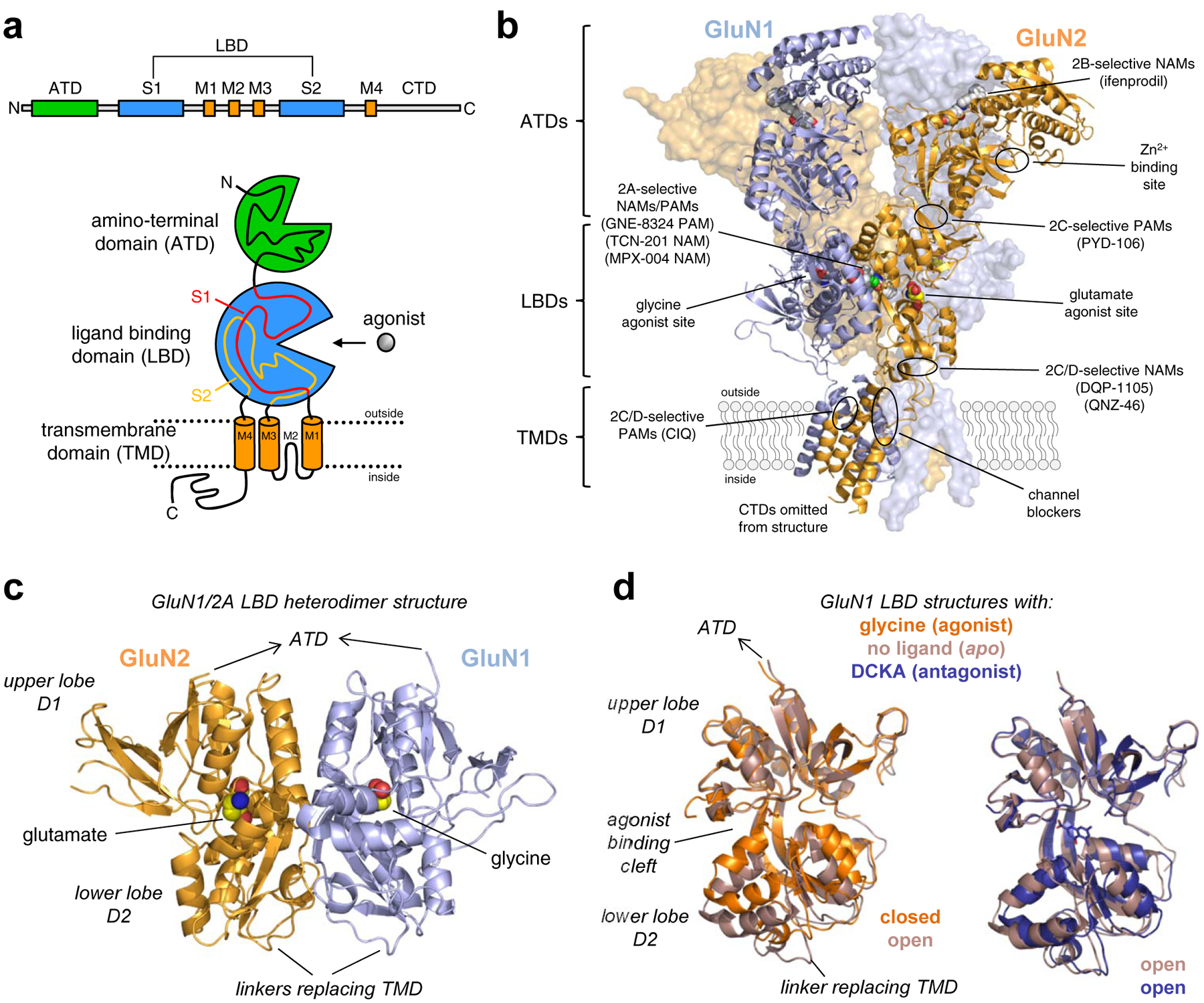

References
-
- Yuzaki M (2009) New (but old) molecules regulating synapse integrity and plasticity: Cbln1 and the delta2 glutamate receptor. Neuroscience 162 (3):633–643 - PubMed
-
- Schmid SM, Hollmann M (2008) To Gate or not to Gate: Are the Delta Subunits in the Glutamate Receptor Family Functional Ion Channels? Mol Neurobiol 37 (2–3):126–141 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
