

doi: 10.1038/nchembio.2493.
Epub 2017 Oct 9.
Stendomycin selectively inhibits TIM23-dependent mitochondrial protein import
Affiliations
- PMID: 28991239
- PMCID: PMC5945950
- DOI: 10.1038/nchembio.2493
Item in Clipboard
Stendomycin selectively inhibits TIM23-dependent mitochondrial protein import
Nat Chem Biol.
2017 Dec.
Abstract
Tim17 and Tim23 are the main subunits of the TIM23 complex, one of the two major essential mitochondrial inner-membrane protein translocon machineries (TIMs). No chemical probes that specifically inhibit TIM23-dependent protein import were known to exist. Here we show that the natural product stendomycin, produced by Streptomyces hygroscopicus, is a potent and specific inhibitor of the TIM23 complex in yeast and mammalian cells. Furthermore, stendomycin-mediated blockage of the TIM23 complex does not alter normal processing of the major regulatory mitophagy kinase PINK1, but TIM23 is required to stabilize PINK1 on the outside of mitochondria to initiate mitophagy upon membrane depolarization.
Figures
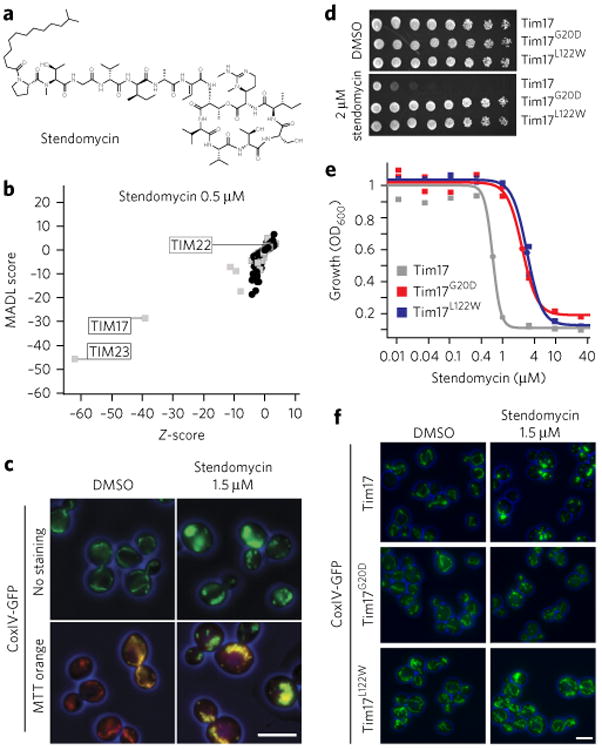
(a) Structure of stendomycin. (b) Haploinsufficiency profiling (HIP) of stendomycin at 0.5 μM. Each mutant strain in the pool is represented by a black circle (nonessential genes) or a gray square (essential genes). The y axis represents strain sensitivity (MADL, mean average distance from log of median), and x axis represents specificity (z-score). Plot shows data calculated from n = 2 technical replicates. (c) Yeast strains carrying a GFP-tagged mitochondrial marker, CoxIV, were treated with 1.5 μM of stendomycin or DMSO for 2 h at 30 °C, stained with mitotracker (MTT) orange, and assessed by fluorescence microscopy (scale bar, 5 μm). (d) Wild-type (Tim17), Tim17G20D and Tim17L122W strains were plated as serial dilutions on solid media (YPD) containing either stendomycin at minimum inhibitory concentration (MIC; 2 μM) or DMSO and incubated at 30 °C for 2 d. (e) Wild-type (Tim17; gray), Tim17G20D (red) and Tim17L122W (blue) strains were grown in duplicate for 18 h at 30 °C in YPD media with 2% DMSO containing a dilution series of stendomycin, and growth was assessed by absorbance at OD600. Dose–response curves were calculated by logistic regression. (f) Wild-type (Tim17), Tim17G20D and Tim17L122W strains were treated with 1.5 μM stendomycin or DMSO for 2 h at 30 °C and assessed for GFP using fluorescence microscopy. Experiments in c,d and f have been replicated twice with similar observations (scale bar, 5 μm).
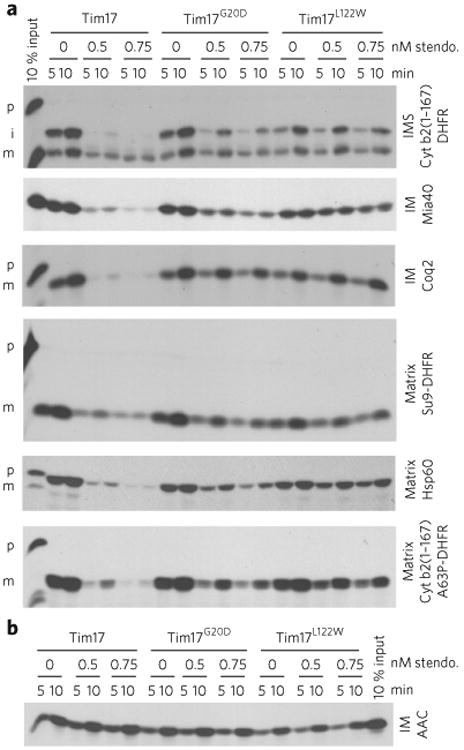
(a) Import assays were performed with radiolabeled TIM23 import pathway substrates into mitochondria isolated from wild-type (Tim17), Tim17G20D and Tim17L122W yeast strains. Time course assays were completed in the presence of 0.5 or 0.75 nM stendomycin (stendo.) or the vehicle control (1% DMSO), and nonimported precursor was removed by trypsin treatment and sodium carbonate extraction. p, precursor; i, intermediate; m, mature. IMS. intermembrane space; IM, inner membrane. (b) As in a, but assessing the TIM22 import pathway substrate AAC. Full gel scans are in Supplementary Information.
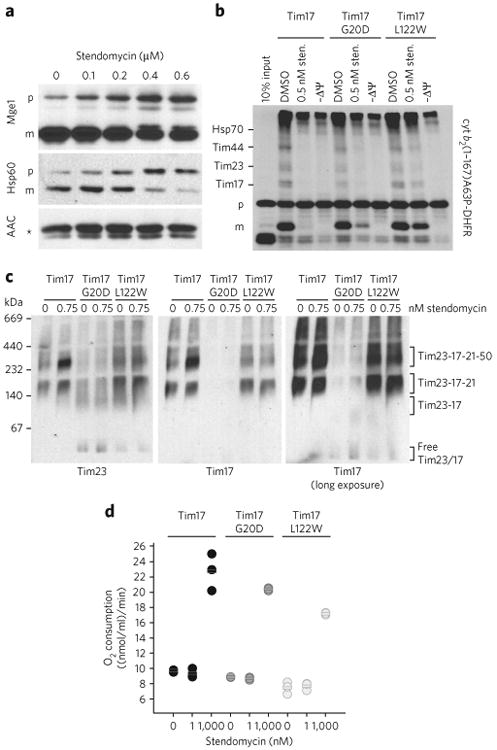
(a) In vivo accumulation of TIM23 complex substrates precursors. Wild-type yeast cells were grown in YPEG media and treated with the indicated concentration of stendomycin for 24 h. The accumulation of Hsp60 and Mge1 precursors was analyzed by immunoblot. The TIM22 complex substrate ACC was used as loading control. p, precursor; m, mature; * marks an unspecific reaction of the antibody. (b) cyt b2(1-167)A63P-DHFR was imported into mitochondria with the addition of 0.5 nM stendomycin or 1% DMSO. The membrane potential was disrupted by CCCP (−ΔΨ). Protein import was arrested by the addition of methotrexate, and cyt b2 (1-167)A63P-DHFR was crosslinked to components of the TIM23 import pathway by DSS. Crosslinked intermediates are indicated. p, precursor; m, mature. Assignment of crosslinked bands was performed as described,. (c) Mitochondria isolated from wild-type, Tim17G20D and Tim17L122W yeast strains were treated with 0.75 nM stendomycin for 15 min, solubilized with digitonin and separated by BN–PAGE. The TIM23 complex was analyzed by immunoblot using antibodies targeting Tim23 and Tim17 (see also Supplementary Fig. 2b). (d) Oxygen consumption of isolated mitochondria from indicated yeast strains was measured with an oxygen electrode. Respiration was initiated by the addition of NADH. 1 or 1,000 nM stendomycin or 1% DMSO was added once a stable respiration had been established. Respiration rates were quantitated based on the DMSO control (see Supplementary Fig. 3 for representative oxygraph plots). Bars represent mean values of n = 3 biological replicates (separate mitochondrial preparations), which are shown as circles (black, wild type; dark grey, Tim17G20D; light grey, Tim17L122W. Full gel scans are in Supplementary Information.
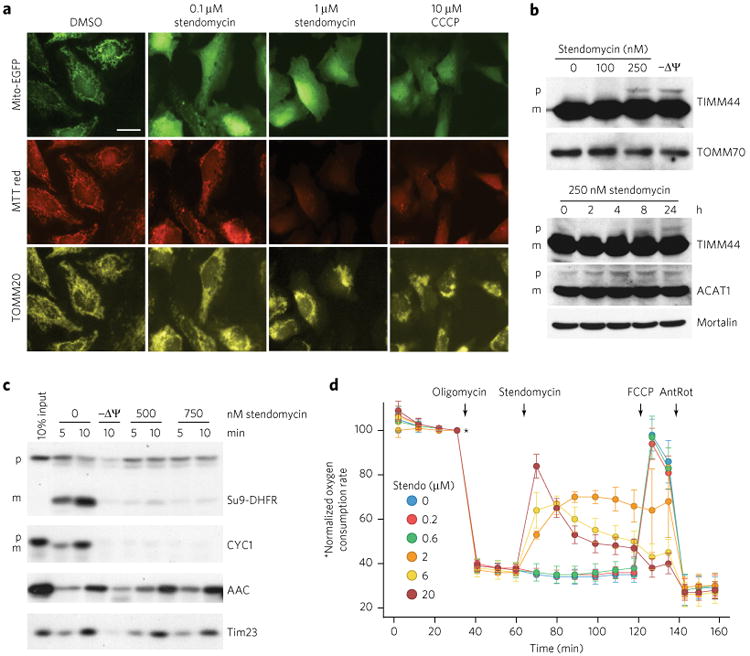
(a) HeLa cells were treated with stendomycin at doses noted or 0.1% DMSO before transfection with MTS-EGFP (mitochondrial targeting sequence of Cox8 fused to EGFP). 10 μM CCCP was used to disrupt the membrane potential (ΔΨ). 24 h after transfection, cells were stained with mitotracker (MTT) red, fixed and visualized for MTS-EGFP, MTT-red and TOMM20. Similar images were obtained in one independent experiment. Scale bar, 10 μM. (b) HeLa cells were treated with the indicated concentrations of stendomycin or with 10 μM CCCP (−ΔΨ) for 24 h (top panel) or with 250 nM stendomycin for the indicated time points (lower panel). Cells were lysed, and total extracts were separated by SDS-PAGE. The accumulation of the TIMM44 and ACAT1 precursors were analyzed by immunoblot. TOMM70 and Mortalin served as loading controls. p, precursor; m, mature. (c) In vitro import assays were performed using mitochondria isolated from HeLa cells and TIM23 and TIM22 import pathways substrates in the presence of 500 or 750 nM stendomycin or control (1% DMSO). Precursors include Su9-DHFR, CYC1, yACC (yeast ACC) and yTim23 (yeast TIM23). p, precursor; m, mature. 10 min time points were quantified and are displayed as percentages in Supplementary Figure 5a. (d) HeLa cells were assessed continuously for oxygen consumption while oligomycin (1 mM), stendomycin (at concentrations noted), FCCP (0.1 μM) and antimycin (1 μM)/rotenone (1 μM; AnRot) were sequentially administered. n = 8 technical replicates; Data represent mean and s.d. This data has been replicated with another passage of HeLa with similar observations. Full gel scans are in Supplementary Information.
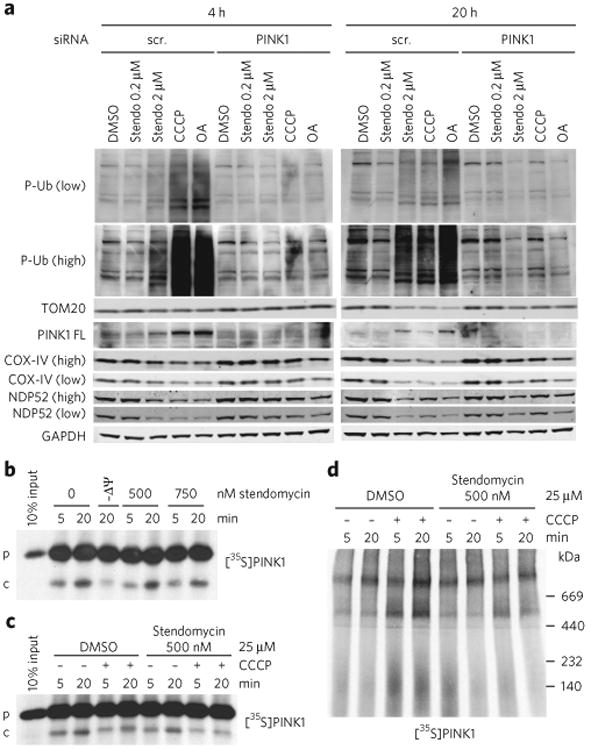
(a) HeLa cells expressing GFP-Parkin were transfected with control (scr, scrambled) or PINK1 siRNAs in the presence of DMSO (0.1%), stendomycin (stendo), CCCP (10 μM) or a combination of oligomycin (100 μM) and antimycin A (100 μM) (OA) for 4 or 20 h. Total cell lysates were analyzed by western blotting as indicated. Similar data was observed in one independent repeat for all markers shown. P-Ub, phospho Ser65-ubiquitin; PINK-FL, full-length PINK1. (b) In vitro import assays were performed as in Figure 4c using PINK1 as substrate. p, precursor; c, PARL-cleaved PINK1. (c) Radiolabeled human PINK1 was in vitro imported into isolated HeLa cell mitochondria in the presence or absence of 500 nM stendomycin for 5 and 20 min. The membrane potential was disrupted in indicated reaction by the addition of 25 μM CCCP. 1% DMSO served as the vehicle control. The membrane insertion of PINK1 was analyzed by alkali extraction for all reactions, which was followed by SDS-PAGE and autoradiography. (d) As in c but with mitochondria solubilized in digitonin and separated by BN-PAGE. PINK1 complexes were visualized by autoradiography. Full gel scans are in Supplementary Information.
Similar articles
-
Architecture of the TIM23 inner mitochondrial translocon and interactions with the matrix import motor.J Biol Chem. 2014 Oct 10;289(41):28689-96. doi: 10.1074/jbc.M114.588152. Epub 2014 Aug 25. J Biol Chem. 2014. PMID: 25157107 Free PMC article.
-
Tim50, a novel component of the TIM23 preprotein translocase of mitochondria.EMBO J. 2003 Feb 17;22(4):816-25. doi: 10.1093/emboj/cdg090. EMBO J. 2003. PMID: 12574118 Free PMC article.
-
Molecular pathway of mitochondrial preprotein import through the TOM-TIM23 supercomplex.Nat Struct Mol Biol. 2023 Dec;30(12):1996-2008. doi: 10.1038/s41594-023-01103-7. Epub 2023 Sep 11. Nat Struct Mol Biol. 2023. PMID: 37696957
-
Protein import into and across the mitochondrial inner membrane: role of the TIM23 and TIM22 translocons.Biochim Biophys Acta. 2002 Sep 2;1592(1):25-34. doi: 10.1016/s0167-4889(02)00261-6. Biochim Biophys Acta. 2002. PMID: 12191765 Review.
-
On the mechanism of preprotein import by the mitochondrial presequence translocase.Biochim Biophys Acta. 2010 Jun;1803(6):732-9. doi: 10.1016/j.bbamcr.2010.01.013. Epub 2010 Jan 25. Biochim Biophys Acta. 2010. PMID: 20100523 Review.
Cited by
-
Disease-Associated Genetic Variation in Human Mitochondrial Protein Import.Am J Hum Genet. 2019 May 2;104(5):784-801. doi: 10.1016/j.ajhg.2019.03.019. Am J Hum Genet. 2019. PMID: 31051112 Free PMC article. Review.
-
Structure-activity relationship of ipglycermide binding to phosphoglycerate mutases.J Biol Chem. 2021 Jan-Jun;296:100628. doi: 10.1016/j.jbc.2021.100628. Epub 2021 Apr 1. J Biol Chem. 2021. PMID: 33812994 Free PMC article.
-
PINK1 import regulation; a fine system to convey mitochondrial stress to the cytosol.BMC Biol. 2018 Jan 10;16(1):2. doi: 10.1186/s12915-017-0470-7. BMC Biol. 2018. PMID: 29325568 Free PMC article. Review.
-
Novel reporter of the PINK1-Parkin mitophagy pathway identifies its damage sensor in the import gate.bioRxiv [Preprint]. 2025 Feb 20:2025.02.19.639160. doi: 10.1101/2025.02.19.639160. bioRxiv. 2025. PMID: 40027798 Free PMC article. Preprint.
-
Visualization of Sirtuin 4 Distribution between Mitochondria and the Nucleus, Based on Bimolecular Fluorescence Self-Complementation.Cells. 2019 Dec 6;8(12):1583. doi: 10.3390/cells8121583. Cells. 2019. PMID: 31817668 Free PMC article.
References
-
- Aoun M, Tiranti V. Mitochondria: a crossroads for lipid metabolism defect in neurodegeneration with brain iron accumulation diseases. Int J Biochem Cell Biol. 2015;63:25–31. - PubMed
-
- Endo T, Yamamoto H, Esaki M. Functional cooperation and separation of translocators in protein import into mitochondria, the double-membrane bounded organelles. J Cell Sci. 2003;116:3259–3267. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
