

Patient-derived xenografts undergo mouse-specific tumor evolution
- PMID: 28991255
- PMCID: PMC5659952
- DOI: 10.1038/ng.3967
Patient-derived xenografts undergo mouse-specific tumor evolution
Abstract
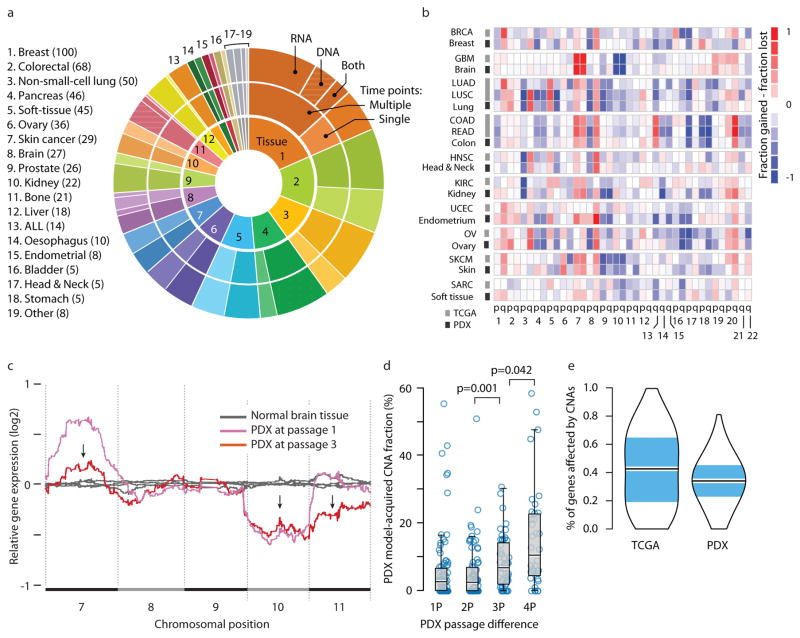
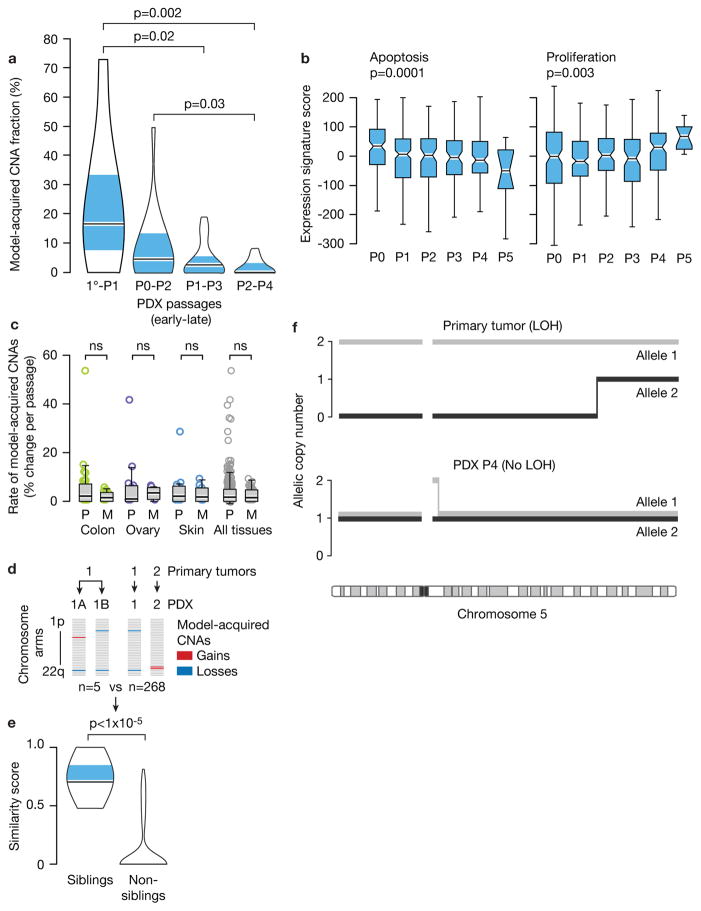
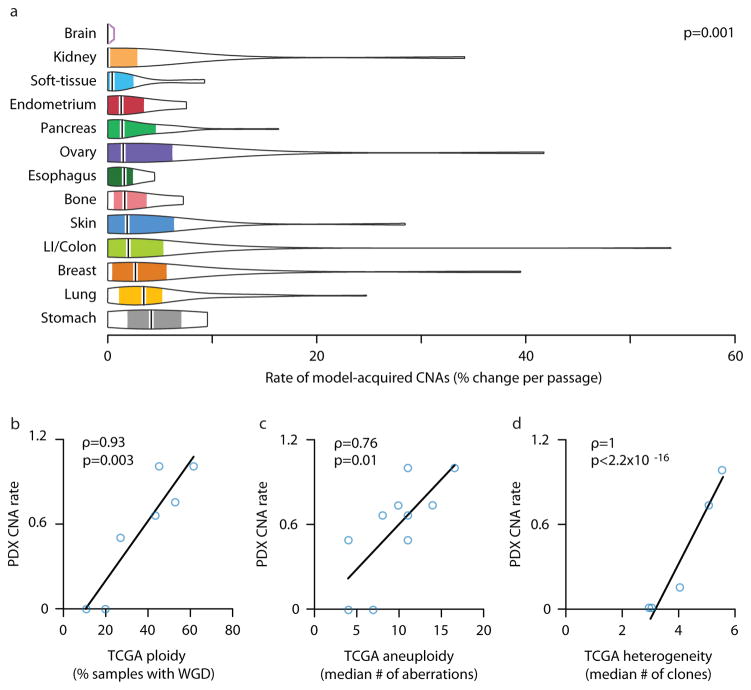
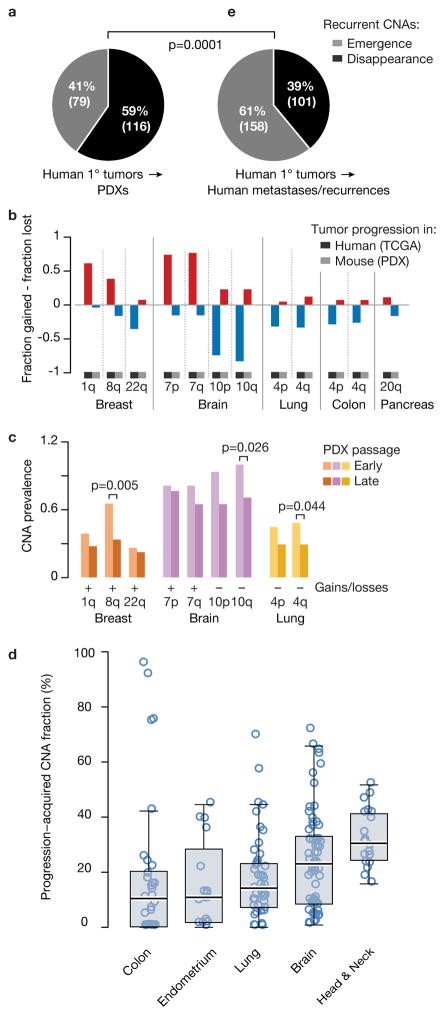
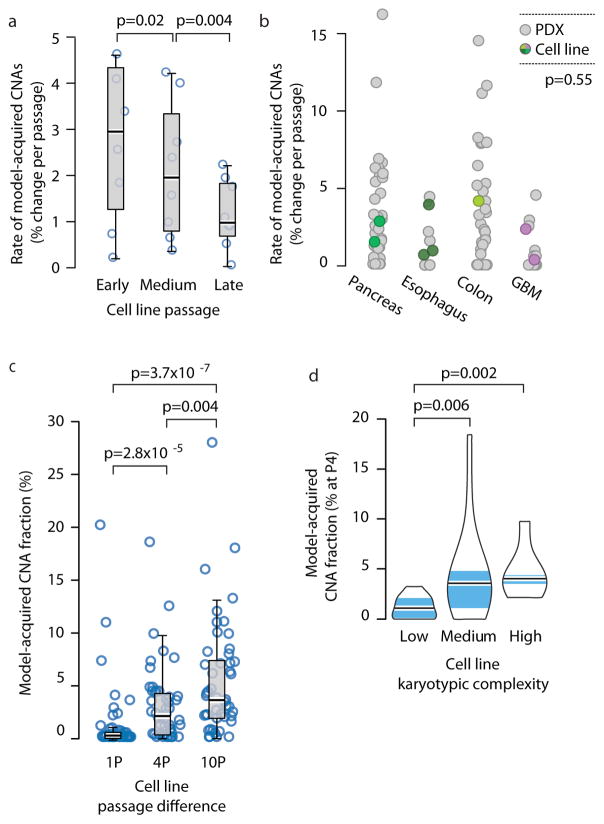
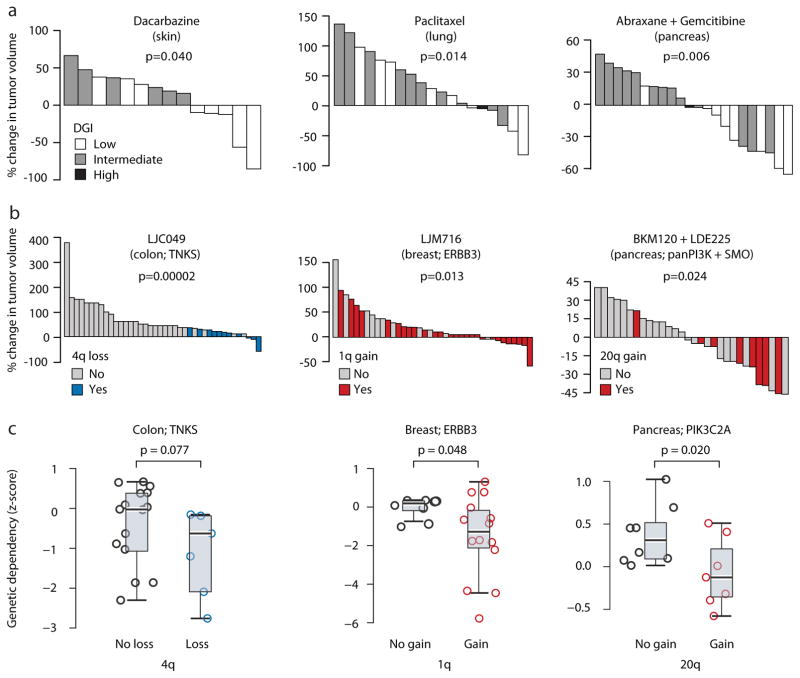
Patient-derived xenografts (PDXs) have become a prominent cancer model system, as they are presumed to faithfully represent the genomic features of primary tumors. Here we monitored the dynamics of copy number alterations (CNAs) in 1,110 PDX samples across 24 cancer types. We observed rapid accumulation of CNAs during PDX passaging, often due to selection of preexisting minor clones. CNA acquisition in PDXs was correlated with the tissue-specific levels of aneuploidy and genetic heterogeneity observed in primary tumors. However, the particular CNAs acquired during PDX passaging differed from those acquired during tumor evolution in patients. Several CNAs recurrently observed in primary tumors gradually disappeared in PDXs, indicating that events undergoing positive selection in humans can become dispensable during propagation in mice. Notably, the genomic stability of PDXs was associated with their response to chemotherapy and targeted drugs. These findings have major implications for PDX-based modeling of human cancer.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Hidalgo M, et al. Patient-derived xenograft models: an emerging platform for translational cancer research. Cancer Discov. 2014;4:998–1013. doi: 10.1158/2159-8290.CD-14-0001. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
