

Epidemiological consequences of immune sensitisation by pre-exposure to vector saliva
- PMID: 28991904
- PMCID: PMC5648264
- DOI: 10.1371/journal.pntd.0005956
Epidemiological consequences of immune sensitisation by pre-exposure to vector saliva
Abstract
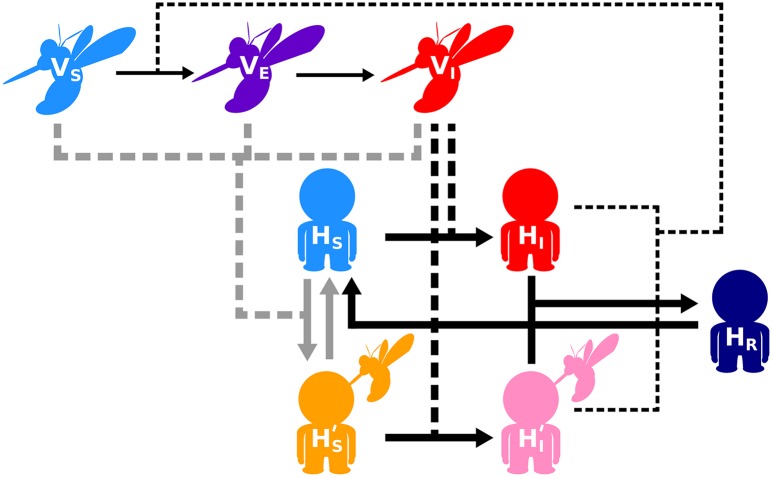
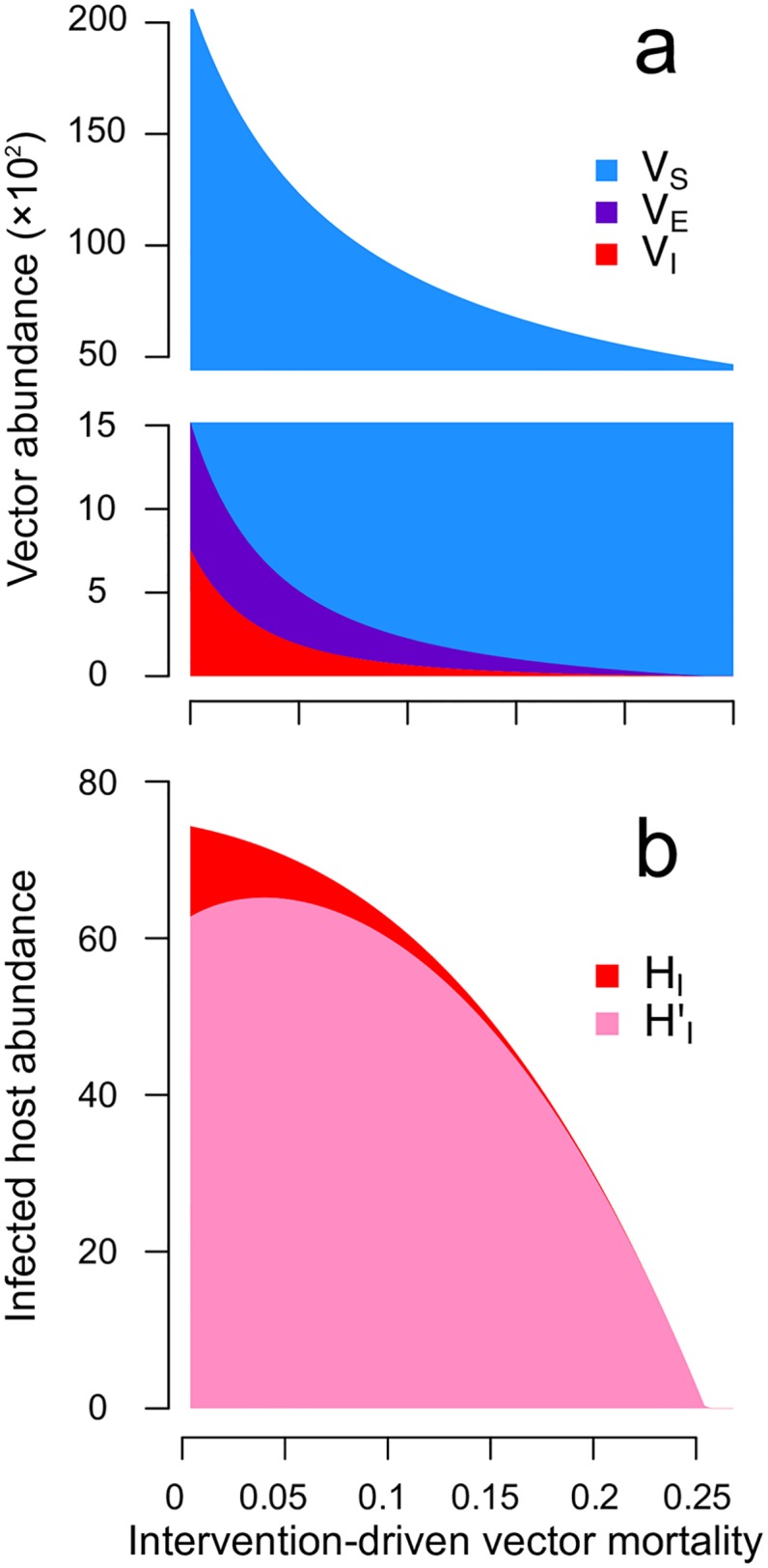
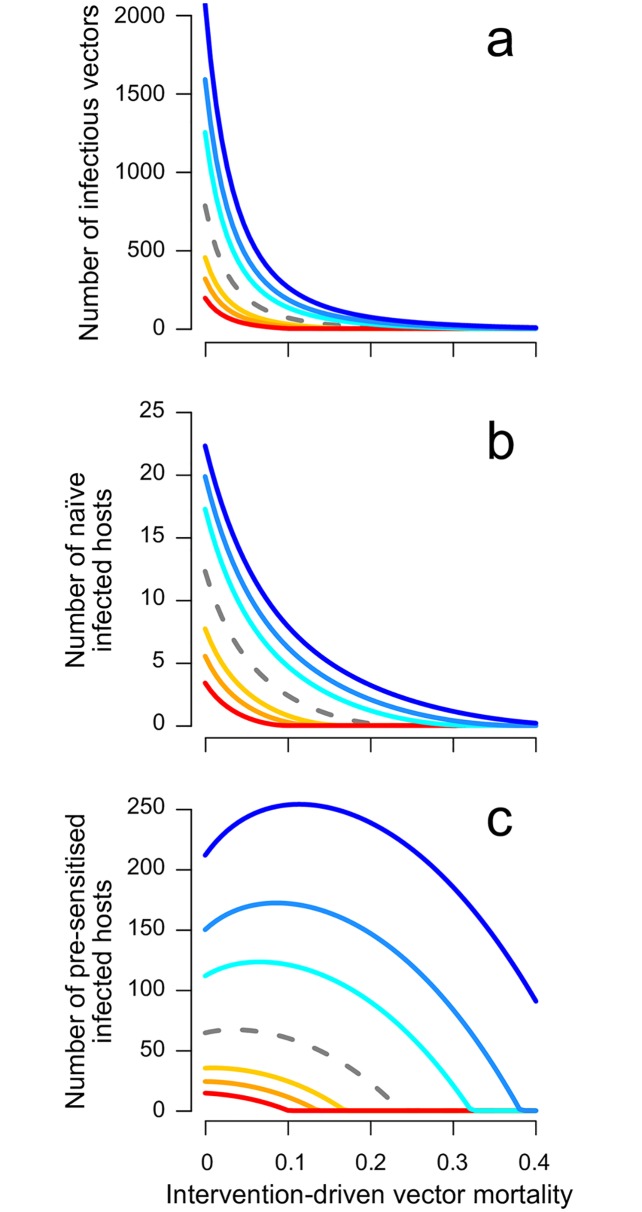
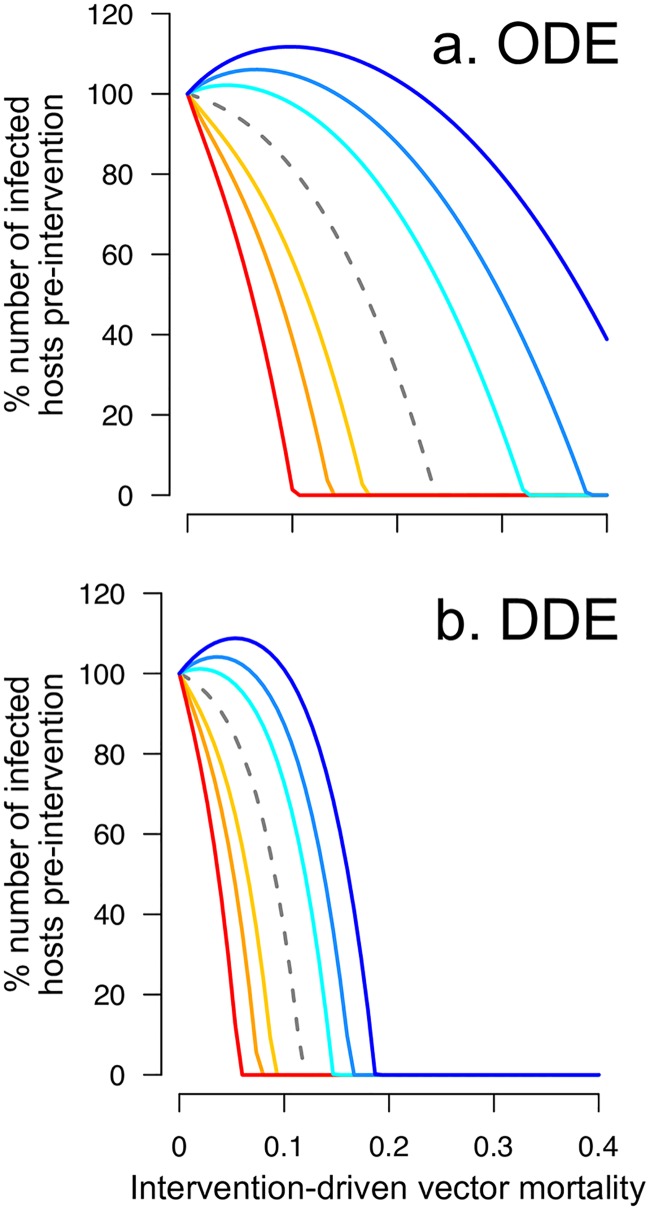
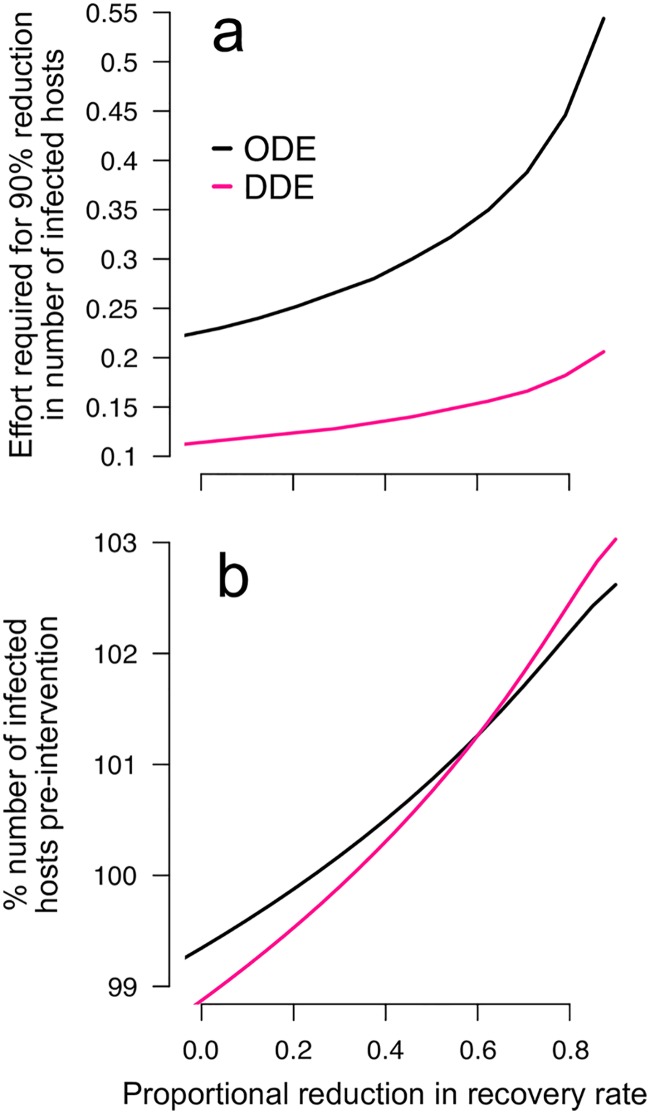
Blood-feeding arthropods-like mosquitoes, sand flies, and ticks-transmit many diseases that impose serious public health and economic burdens. When a blood-feeding arthropod bites a mammal, it injects saliva containing immunogenic compounds that facilitate feeding. Evidence from Leishmania, Plasmodium and arboviral infections suggests that the immune responses elicited by pre-exposure to arthropod saliva can alter disease progression if the host later becomes infected. Such pre-sensitisation of host immunity has been reported to both exacerbate and limit infection symptoms, depending on the system in question, with potential implications for recovery. To explore if and how immune pre-sensitisation alters the effects of vector control, we develop a general model of vector-borne disease. We show that the abundance of pre-sensitised infected hosts should increase when control efforts moderately increase vector mortality rates. If immune pre-sensitisation leads to more rapid clearance of infection, increasing vector mortality rates may achieve greater than expected disease control. However, when immune pre-sensitisation prolongs the duration of infection, e.g., through mildly symptomatic cases for which treatment is unlikely to be sought, vector control can actually increase the total number of infected hosts. The rising infections may go unnoticed unless active surveillance methods are used to detect such sub-clinical individuals, who could provide long-lasting reservoirs for transmission and suffer long-term health consequences of those sub-clinical infections. Sensitivity analysis suggests that these negative consequences could be mitigated through integrated vector management. While the effect of saliva pre-exposure on acute symptoms is well-studied for leishmaniasis, the immunological and clinical consequences are largely uncharted for other vector-parasite-host combinations. We find a large range of plausible epidemiological outcomes, positive and negative for public health, underscoring the need to quantify how immune pre-sensitisation modulates recovery and transmission rates in vector-borne diseases.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Modulation of host immunity by haematophagous arthropods.Ann Trop Med Parasitol. 2001 Dec;95(8):755-71. doi: 10.1080/0003498012011118. Ann Trop Med Parasitol. 2001. PMID: 11784430 Review.
-
Progress toward molecular characterization of ectoparasite modulation of host immunity.Vet Parasitol. 2001 Nov 22;101(3-4):275-87. doi: 10.1016/s0304-4017(01)00556-8. Vet Parasitol. 2001. PMID: 11707302 Review.
-
The immunomodulatory factors of arthropod saliva and the potential for these factors to serve as vaccine targets to prevent pathogen transmission.Parasite Immunol. 2006 Apr;28(4):131-41. doi: 10.1111/j.1365-3024.2006.00807.x. Parasite Immunol. 2006. PMID: 16542315 Review.
-
Immunological consequences of arthropod vector-derived salivary factors.Eur J Immunol. 2011 Dec;41(12):3396-400. doi: 10.1002/eji.201190075. Eur J Immunol. 2011. PMID: 22125007
-
Immune responses to arthropods and their products.Annu Rev Entomol. 1982;27:21-48. doi: 10.1146/annurev.en.27.010182.000321. Annu Rev Entomol. 1982. PMID: 6123290 Review. No abstract available.
Cited by
-
Leishmaniasis in deployed military populations: A systematic review and meta-analysis.PLoS Negl Trop Dis. 2025 Mar 10;19(3):e0012680. doi: 10.1371/journal.pntd.0012680. eCollection 2025 Mar. PLoS Negl Trop Dis. 2025. PMID: 40063644 Free PMC article.
-
Anopheles salivary antigens as serological biomarkers of vector exposure and malaria transmission: A systematic review with multilevel modelling.Elife. 2021 Dec 23;10:e73080. doi: 10.7554/eLife.73080. Elife. 2021. PMID: 34939933 Free PMC article.
-
Estimating time-dependent contact: a multi-strain epidemiological model of SARS-CoV-2 on the island of Ireland.Glob Epidemiol. 2023 Dec;5:100111. doi: 10.1016/j.gloepi.2023.100111. Epub 2023 Apr 28. Glob Epidemiol. 2023. PMID: 37162815 Free PMC article.
References
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
