

Timing avian long-distance migration: from internal clock mechanisms to global flights
- PMID: 28993496
- PMCID: PMC5647279
- DOI: 10.1098/rstb.2016.0252
Timing avian long-distance migration: from internal clock mechanisms to global flights
Abstract
Migratory birds regularly perform impressive long-distance flights, which are timed relative to the anticipated environmental resources at destination areas that can be several thousand kilometres away. Timely migration requires diverse strategies and adaptations that involve an intricate interplay between internal clock mechanisms and environmental conditions across the annual cycle. Here we review what challenges birds face during long migrations to keep track of time as they exploit geographically distant resources that may vary in availability and predictability, and summarize the clock mechanisms that enable them to succeed. We examine the following challenges: departing in time for spring and autumn migration, in anticipation of future environmental conditions; using clocks on the move, for example for orientation, navigation and stopover; strategies of adhering to, or adjusting, the time programme while fitting their activities into an annual cycle; and keeping pace with a world of rapidly changing environments. We then elaborate these themes by case studies representing long-distance migrating birds with different annual movement patterns and associated adaptations of their circannual programmes. We discuss the current knowledge on how endogenous migration programmes interact with external information across the annual cycle, how components of annual cycle programmes encode topography and range expansions, and how fitness may be affected when mismatches between timing and environmental conditions occur. Lastly, we outline open questions and propose future research directions.This article is part of the themed issue 'Wild clocks: integrating chronobiology and ecology to understand timekeeping in free-living animals'.
Keywords: circannual programmes; clock; environment; migration strategies; orientation; photoperiod.
© 2017 The Author(s).
Conflict of interest statement
We have no competing interests.
Figures
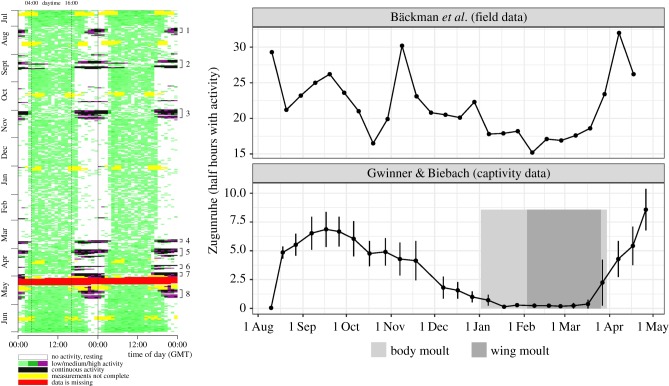
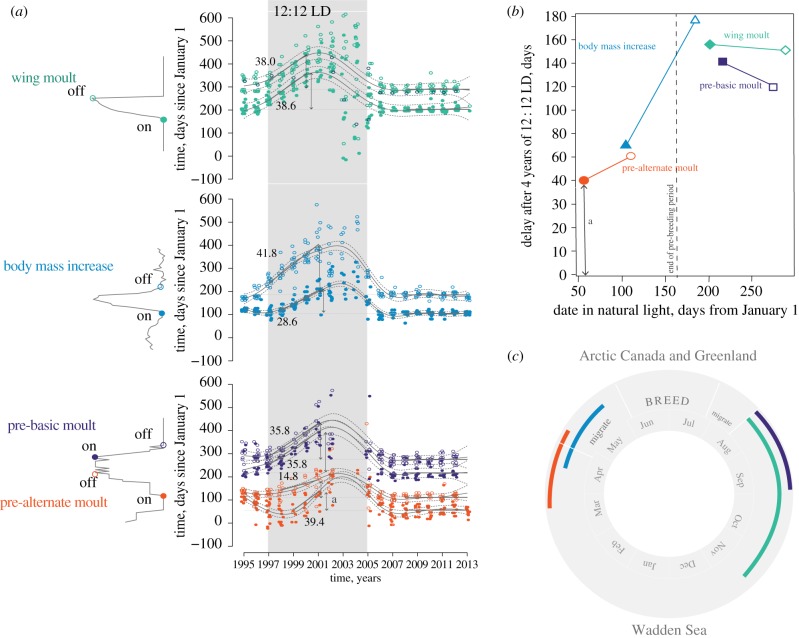
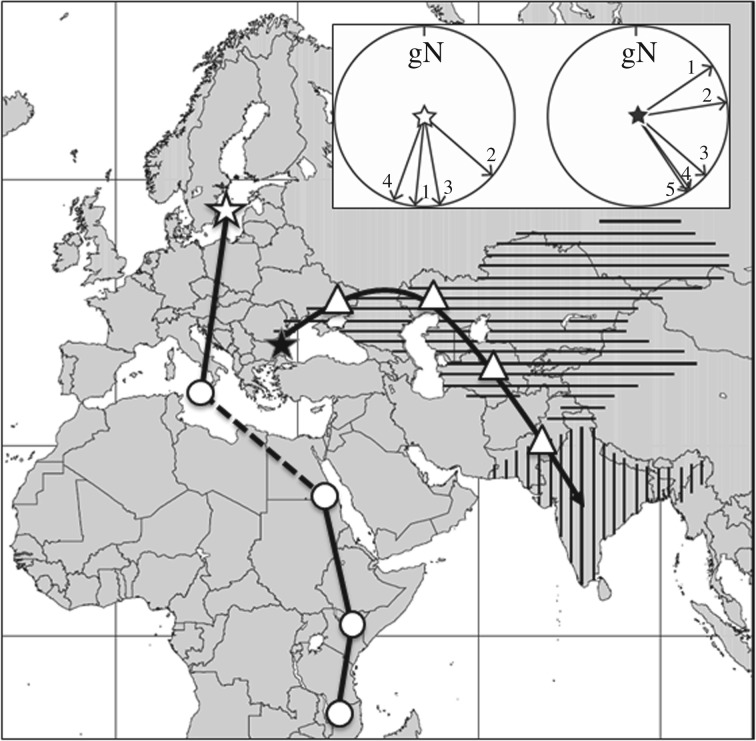
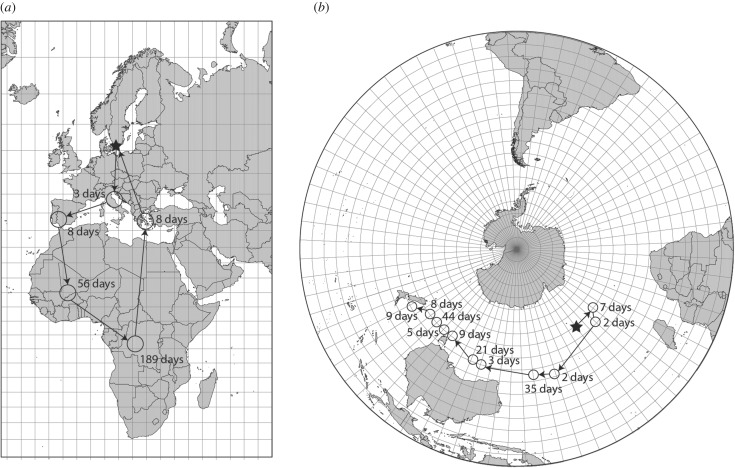
References
-
- Rowan W. 1926. On photoperiodism, reproductive periodicity, and the annual migrations of birds and certain fishes. Proc. Boston Soc. Nat. Hist. 38, 147–189.
-
- Aschoff J. 1955. Jahresperiodik der Fortpflanzung bei Warmblütern. Stud. Gen. 8, 742–776.
-
- Gwinner E. 1996. Circannual clocks in avian reproduction and migration. Ibis 138, 47–63. ( 10.1111/j.1474-919X.1996.tb04312.x) - DOI
-
- Berthold P. 1996. Control of bird migration. Berlin, Germany: Springer.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources