

Calcium and electrical dynamics in lymphatic endothelium
- PMID: 28994159
- PMCID: PMC5730853
- DOI: 10.1113/JP274842
Calcium and electrical dynamics in lymphatic endothelium
Abstract
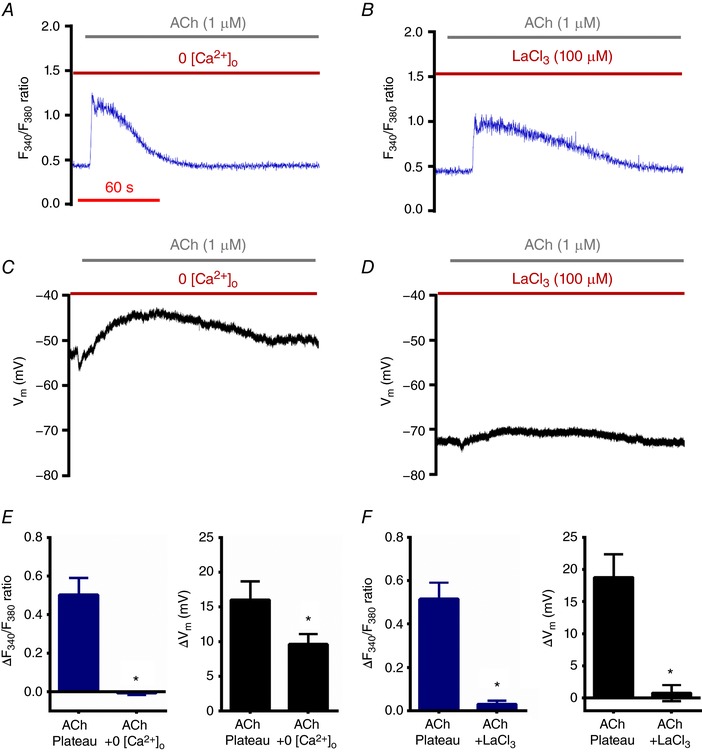
Key points: Endothelial cell function in resistance arteries integrates Ca2+ signalling with hyperpolarization to promote relaxation of smooth muscle cells and increase tissue blood flow. Whether complementary signalling occurs in lymphatic endothelium is unknown. Intracellular calcium and membrane potential were evaluated in endothelial cell tubes freshly isolated from mouse collecting lymphatic vessels of the popliteal fossa. Resting membrane potential measured using intracellular microelectrodes averaged ∼-70 mV. Stimulation of lymphatic endothelium by acetylcholine or a TRPV4 channel agonist increased intracellular Ca2+ with robust depolarization. Findings from Trpv4-/- mice and with computational modelling suggest that the initial mobilization of intracellular Ca2+ leads to influx of Ca2+ and Na+ through TRPV4 channels to evoke depolarization. Lymphatic endothelial cells lack the Ca2+ -activated K+ channels present in arterial endothelium to generate endothelium-derived hyperpolarization. Absence of this signalling pathway with effective depolarization may promote rapid conduction of contraction along lymphatic muscle during lymph propulsion.
Abstract: Subsequent to a rise in intracellular Ca2+ ([Ca2+ ]i ), hyperpolarization of the endothelium coordinates vascular smooth muscle relaxation along resistance arteries during blood flow control. In the lymphatic vasculature, collecting vessels generate rapid contractions coordinated along lymphangions to propel lymph, but the underlying signalling pathways are unknown. We tested the hypothesis that lymphatic endothelial cells (LECs) exhibit Ca2+ and electrical signalling properties that facilitate lymph propulsion. To study electrical and intracellular Ca2+ signalling dynamics in lymphatic endothelium, we excised collecting lymphatic vessels from the popliteal fossa of mice and removed their muscle cells to isolate intact LEC tubes (LECTs). Intracellular recording revealed a resting membrane potential of ∼-70 mV. Acetylcholine (ACh) increased [Ca2+ ]i with a time course similar to that observed in endothelium of resistance arteries (i.e. rapid initial peak with a sustained 'plateau'). In striking contrast to the endothelium-derived hyperpolarization (EDH) characteristic of arteries, LECs depolarized (>15 mV) to either ACh or TRPV4 channel activation. This depolarization was facilitated by the absence of Ca2+ -activated K+ (KCa ) channels as confirmed with PCR, persisted in the absence of extracellular Ca2+ , was abolished by LaCl3 and was attenuated ∼70% in LECTs from Trpv4-/- mice. Computational modelling of ion fluxes in LECs indicated that omitting K+ channels supports our experimental results. These findings reveal novel signalling events in LECs, which are devoid of the KCa activity abundant in arterial endothelium. Absence of EDH with effective depolarization of LECs may promote the rapid conduction of contraction waves along lymphatic muscle during lymph propulsion.
Keywords: TRP channels; calcium-activated K+ channel; endothelium-derived hyperpolarizing factor; mathematical model.
© 2017 The Authors. The Journal of Physiology © 2017 The Physiological Society.
Figures









Comment in
-
Endothelial tubes: another window into lymphatic function.J Physiol. 2017 Dec 15;595(24):7267-7268. doi: 10.1113/JP275323. Epub 2017 Nov 21. J Physiol. 2017. PMID: 29063612 Free PMC article. No abstract available.
References
-
- Aspelund A, Robciuc MR, Karaman S, Makinen T & Alitalo K (2016). Lymphatic system in cardiovascular medicine. Circ Res 118, 515–530. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 HL120867/HL/NHLBI NIH HHS/United States
- R01 HL122578/HL/NHLBI NIH HHS/United States
- F32 HL110701/HL/NHLBI NIH HHS/United States
- U01 HL123420/HL/NHLBI NIH HHS/United States
- R00 AG047198/AG/NIA NIH HHS/United States
- R00 HL124142/HL/NHLBI NIH HHS/United States
- K99 AG047198/AG/NIA NIH HHS/United States
- R01 HL094269/HL/NHLBI NIH HHS/United States
- K99 HL124142/HL/NHLBI NIH HHS/United States
- R01 HL041026/HL/NHLBI NIH HHS/United States
- R01 HL125608/HL/NHLBI NIH HHS/United States
- R37 HL041026/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous