

Targeted mutagenesis in a human-parasitic nematode
- PMID: 29016680
- PMCID: PMC5650185
- DOI: 10.1371/journal.ppat.1006675
Targeted mutagenesis in a human-parasitic nematode
Abstract
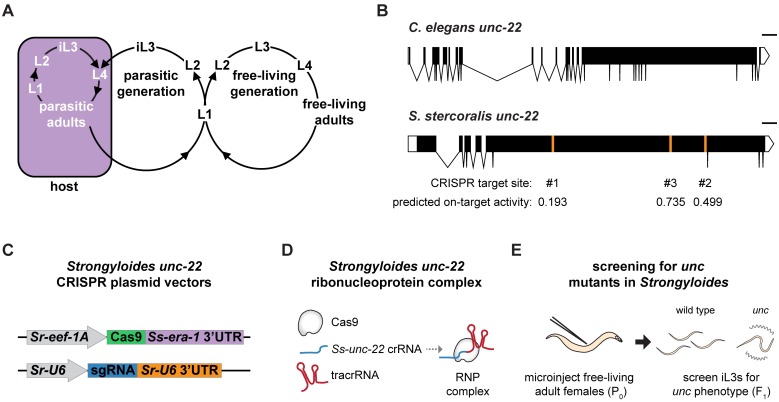
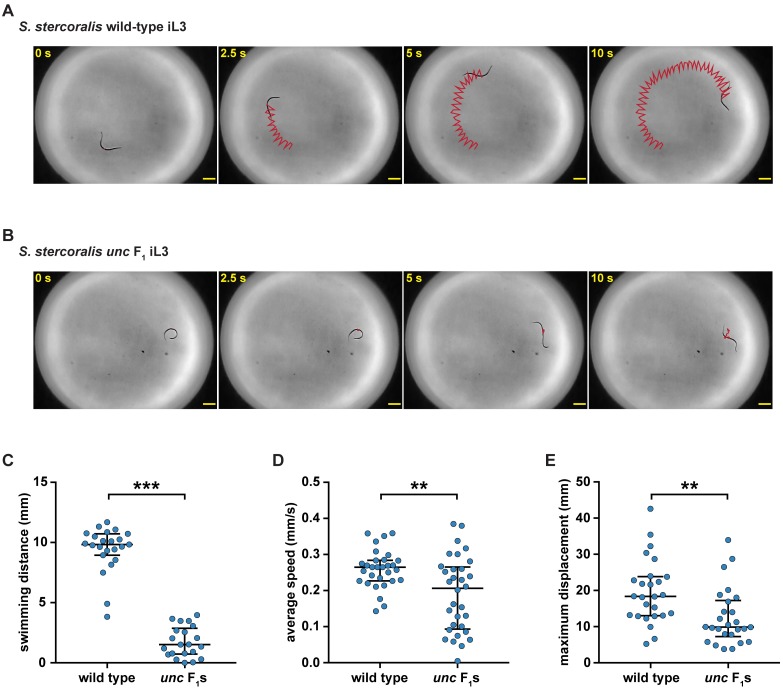
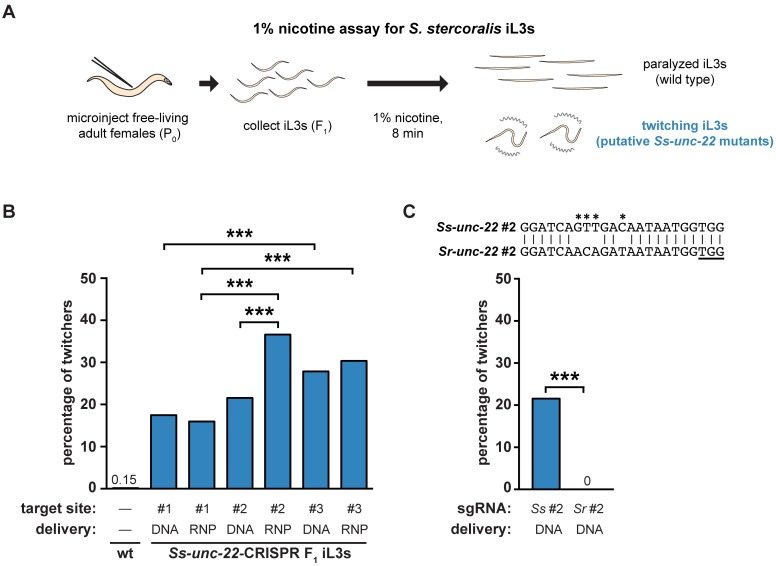
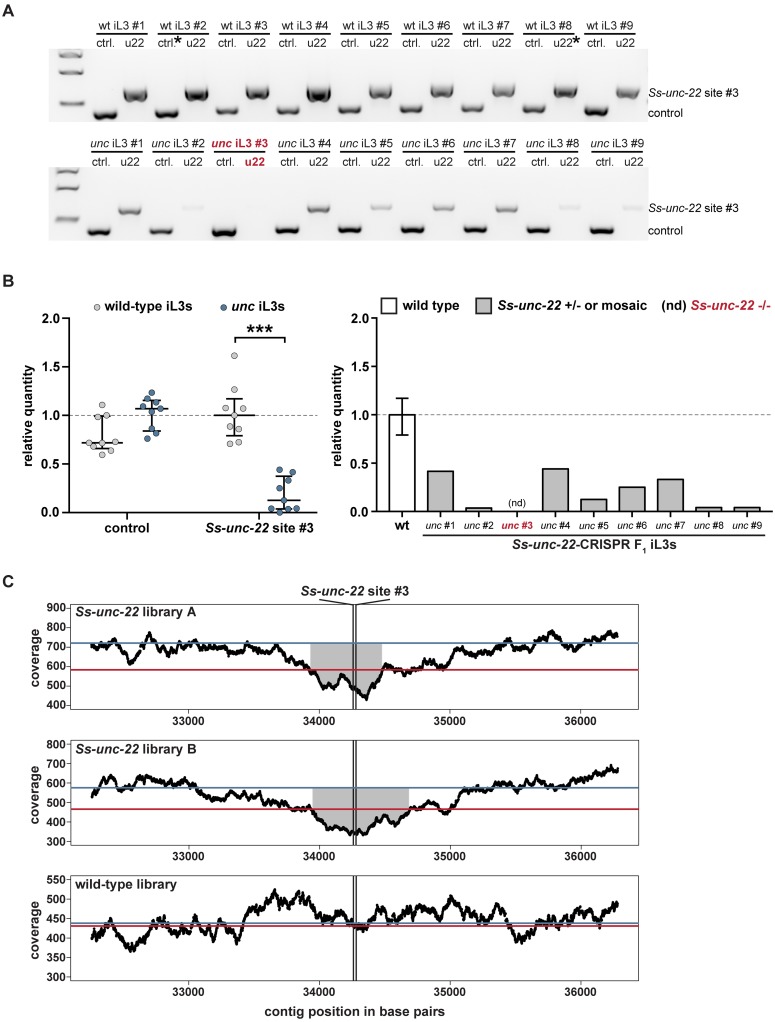
Parasitic nematodes infect over 1 billion people worldwide and cause some of the most common neglected tropical diseases. Despite their prevalence, our understanding of the biology of parasitic nematodes has been limited by the lack of tools for genetic intervention. In particular, it has not yet been possible to generate targeted gene disruptions and mutant phenotypes in any parasitic nematode. Here, we report the development of a method for introducing CRISPR-Cas9-mediated gene disruptions in the human-parasitic threadworm Strongyloides stercoralis. We disrupted the S. stercoralis twitchin gene unc-22, resulting in nematodes with severe motility defects. Ss-unc-22 mutations were resolved by homology-directed repair when a repair template was provided. Omission of a repair template resulted in deletions at the target locus. Ss-unc-22 mutations were heritable; we passed Ss-unc-22 mutants through a host and successfully recovered mutant progeny. Using a similar approach, we also disrupted the unc-22 gene of the rat-parasitic nematode Strongyloides ratti. Our results demonstrate the applicability of CRISPR-Cas9 to parasitic nematodes, and thereby enable future studies of gene function in these medically relevant but previously genetically intractable parasites.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
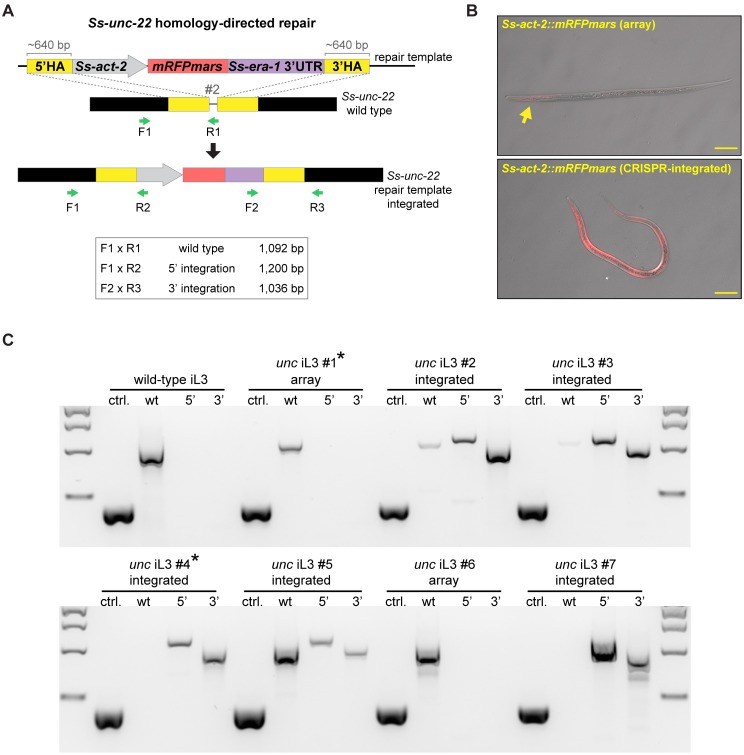
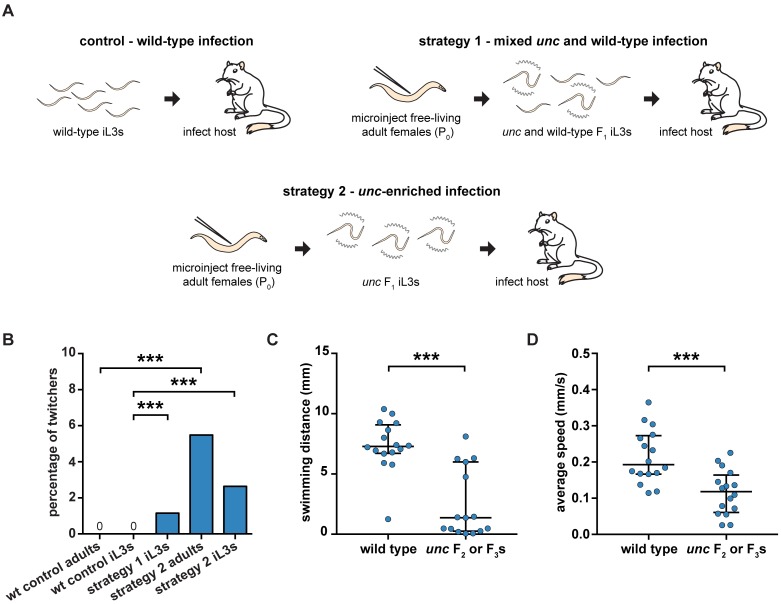
Comment in
-
Spotlight on CRISPR in Strongyloides Parasitic Nematodes.Trends Parasitol. 2018 Jan;34(1):6-9. doi: 10.1016/j.pt.2017.11.005. Epub 2017 Nov 23. Trends Parasitol. 2018. PMID: 29174224 Free PMC article.
Similar articles
-
Generating Transgenics and Knockouts in Strongyloides Species by Microinjection.J Vis Exp. 2021 Oct 7;(176):10.3791/63023. doi: 10.3791/63023. J Vis Exp. 2021. PMID: 34694289 Free PMC article.
-
Terror in the dirt: Sensory determinants of host seeking in soil-transmitted mammalian-parasitic nematodes.Int J Parasitol Drugs Drug Resist. 2018 Dec;8(3):496-510. doi: 10.1016/j.ijpddr.2018.10.008. Epub 2018 Oct 26. Int J Parasitol Drugs Drug Resist. 2018. PMID: 30396862 Free PMC article. Review.
-
Transgenesis in the parasitic nematode Strongyloides ratti.Mol Biochem Parasitol. 2011 Oct;179(2):114-9. doi: 10.1016/j.molbiopara.2011.06.002. Epub 2011 Jun 23. Mol Biochem Parasitol. 2011. PMID: 21723330 Free PMC article.
-
Transposon-mediated chromosomal integration of transgenes in the parasitic nematode Strongyloides ratti and establishment of stable transgenic lines.PLoS Pathog. 2012;8(8):e1002871. doi: 10.1371/journal.ppat.1002871. Epub 2012 Aug 9. PLoS Pathog. 2012. PMID: 22912584 Free PMC article.
-
Invade or die: behaviours and biochemical mechanisms that drive skin penetration in Strongyloides and other skin-penetrating nematodes.Philos Trans R Soc Lond B Biol Sci. 2024 Jan 15;379(1894):20220434. doi: 10.1098/rstb.2022.0434. Epub 2023 Nov 27. Philos Trans R Soc Lond B Biol Sci. 2024. PMID: 38008119 Free PMC article. Review.
Cited by
-
Clustered Regularly Interspaced Short Palindromic Repeats/ CRISPR associated protein 9-mediated editing of Schistosoma mansoni genes: Identifying genes for immunologically potent drug and vaccine development.Rev Soc Bras Med Trop. 2022 Aug 12;55:e0131. doi: 10.1590/0037-8682-0131-2022. eCollection 2022. Rev Soc Bras Med Trop. 2022. PMID: 35976333 Free PMC article. Review.
-
Exposure to dexamethasone modifies transcriptomic responses of free-living stages of Strongyloides stercoralis.PLoS One. 2021 Jun 28;16(6):e0253701. doi: 10.1371/journal.pone.0253701. eCollection 2021. PLoS One. 2021. PMID: 34181669 Free PMC article.
-
Dopamine signaling drives skin invasion by human-infective nematodes.Nat Commun. 2025 Aug 13;16(1):7246. doi: 10.1038/s41467-025-62517-z. Nat Commun. 2025. PMID: 40804046 Free PMC article.
-
Carbon dioxide shapes parasite-host interactions in a human-infective nematode.bioRxiv [Preprint]. 2024 Mar 31:2024.03.28.587273. doi: 10.1101/2024.03.28.587273. bioRxiv. 2024. Update in: Curr Biol. 2025 Jan 20;35(2):277-286.e6. doi: 10.1016/j.cub.2024.11.036. PMID: 38585813 Free PMC article. Updated. Preprint.
-
Caenorhabditis elegans in anthelmintic research - Old model, new perspectives.Int J Parasitol Drugs Drug Resist. 2020 Dec;14:237-248. doi: 10.1016/j.ijpddr.2020.09.005. Epub 2020 Oct 2. Int J Parasitol Drugs Drug Resist. 2020. PMID: 33249235 Free PMC article. Review.
References
-
- Pullan RL, Smith JL, Jasrasaria R, Brooker SJ. Global numbers of infection and disease burden of soil transmitted helminth infections in 2010. Parasit Vectors. 2014;7:37 doi: 10.1186/1756-3305-7-37 - DOI - PMC - PubMed
-
- Boatin BA, Basanez MG, Prichard RK, Awadzi K, Barakat RM, Garcia HH, et al. A research agenda for helminth diseases of humans: towards control and elimination. PLoS Negl Trop Dis. 2012;6(4):e1547 doi: 10.1371/journal.pntd.0001547 - DOI - PMC - PubMed
-
- Ward JD. Rendering the intractable more tractable: tools from Caenorhabditis elegans ripe for import into parasitic nematodes. Genetics. 2015;201(4):1279–94. doi: 10.1534/genetics.115.182717 - DOI - PMC - PubMed
-
- Guo L, Chang Z, Dieterich C, Streit A. A protocol for chemical mutagenesis in Strongyloides ratti. Exp Parasitol. 2015;158:2–7. doi: 10.1016/j.exppara.2015.03.001 - DOI - PubMed
-
- Doudna JA, Charpentier E. The new frontier of genome engineering with CRISPR-Cas9. Science. 2014;346(6213):1258096 doi: 10.1126/science.1258096 - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
