

Potassium channel dysfunction underlies Purkinje neuron spiking abnormalities in spinocerebellar ataxia type 2
- PMID: 29016852
- PMCID: PMC5886219
- DOI: 10.1093/hmg/ddx281
Potassium channel dysfunction underlies Purkinje neuron spiking abnormalities in spinocerebellar ataxia type 2
Abstract
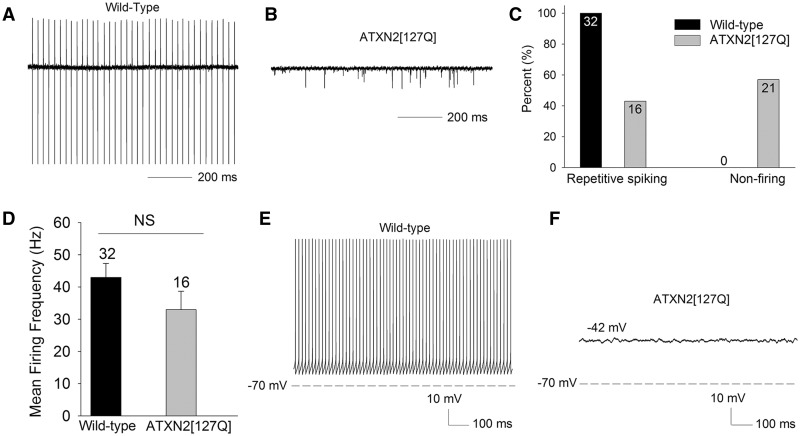
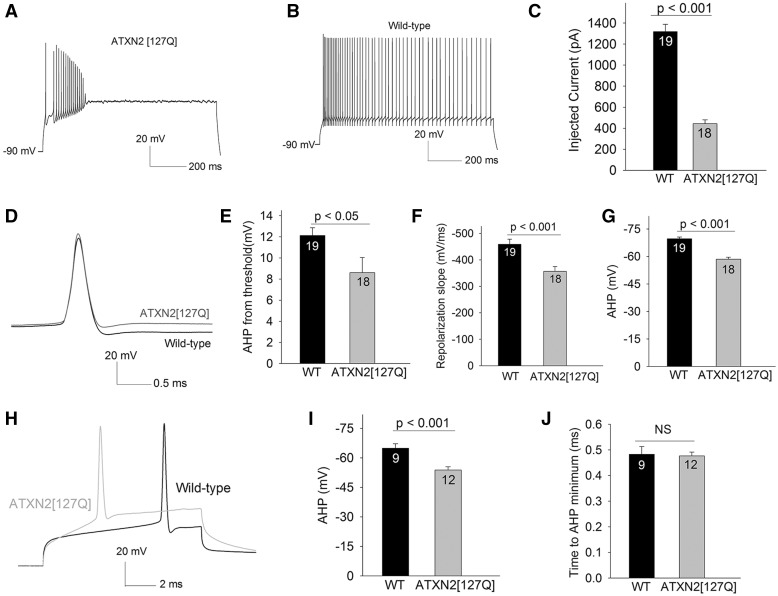
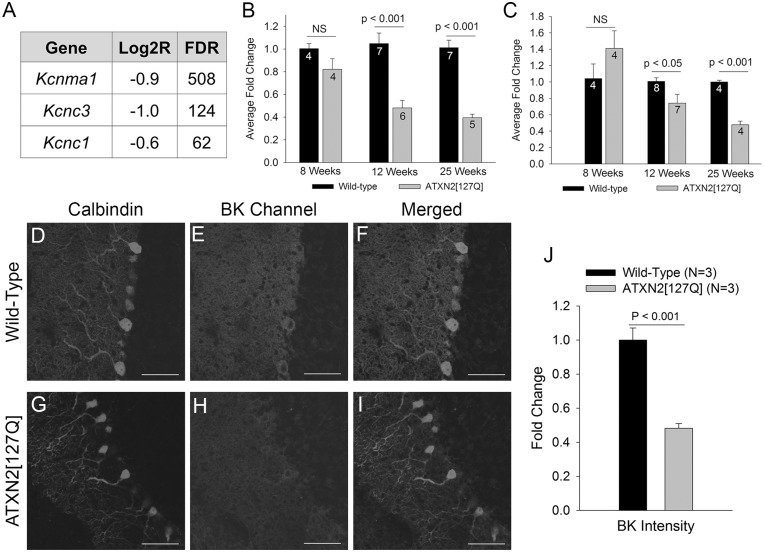
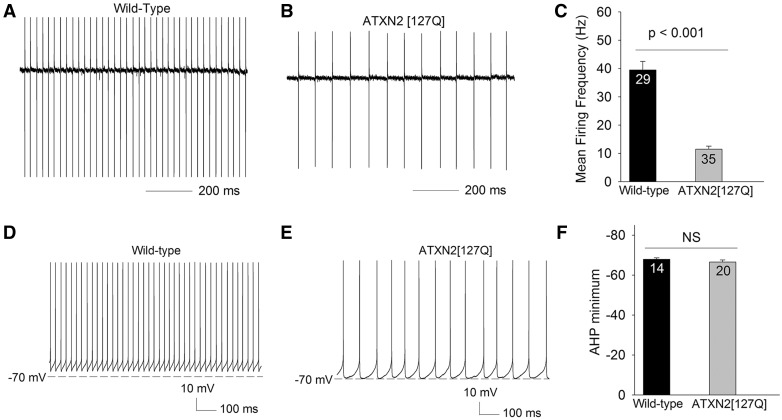
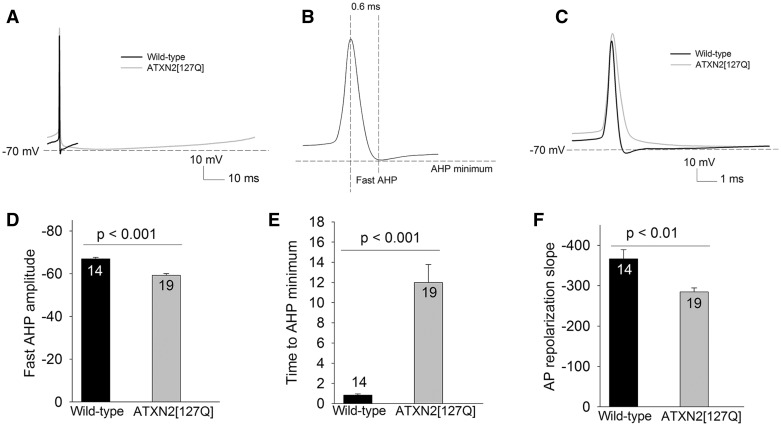
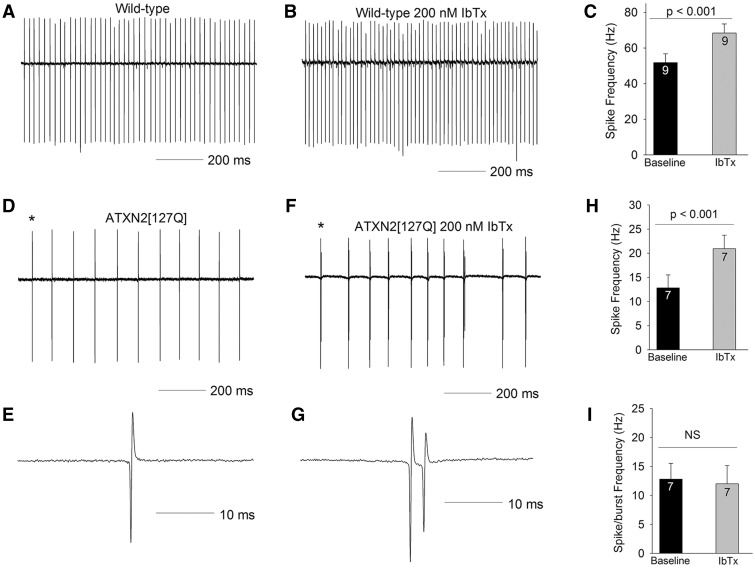
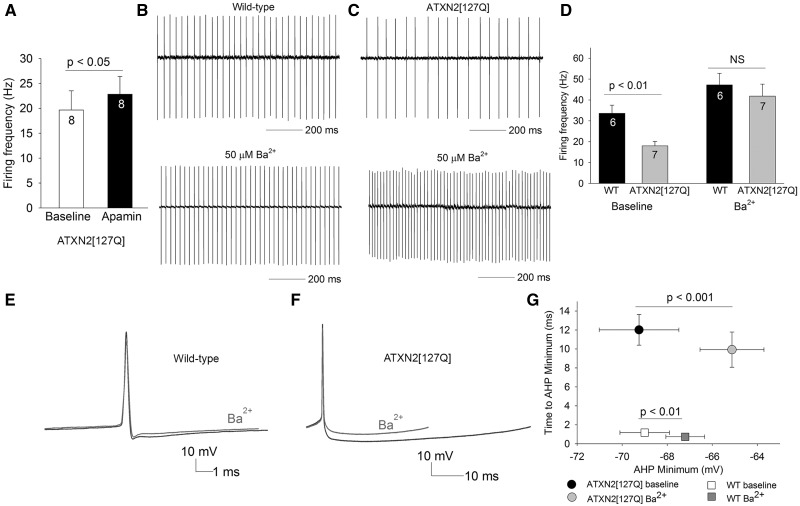
Alterations in Purkinje neuron firing often accompany ataxia, but the molecular basis for these changes is poorly understood. In a mouse model of spinocerebellar ataxia type 2 (SCA2), a progressive reduction in Purkinje neuron firing frequency accompanies cell atrophy. We investigated the basis for altered Purkinje neuron firing in SCA2. A reduction in the expression of large-conductance calcium-activated potassium (BK) channels and Kv3.3 voltage-gated potassium channels accompanies the inability of Purkinje neurons early in disease to maintain repetitive spiking. In association with prominent Purkinje neuron atrophy, repetitive spiking is restored, although at a greatly reduced firing frequency. In spite of a continued impairment in spike repolarization and a persistently reduced BK channel mediated afterhyperpolarization (AHP), repetitive spiking is maintained, through the increased activity of barium-sensitive potassium channels, most consistent with inwardly rectifying potassium (Kir) channels. Increased activity of Kir channels results in the generation of a novel AHP not seen in wild-type Purkinje neurons that also accounts for the reduced firing frequency late in disease. Homeostatic changes in Purkinje neuron morphology that help to preserve repetitive spiking can also therefore have deleterious consequences for spike frequency. These results suggest that the basis for spiking abnormalities in SCA2 differ depending on disease stage, and interventions targeted towards correcting potassium channel dysfunction in ataxia need to be tailored to the specific stage in the degenerative process.
© The Author 2017. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures
References
-
- Hausser M., Clark B.A. (1997) Tonic synaptic inhibition modulates neuronal output pattern and spatiotemporal synaptic integration. Neuron, 19, 665–678. - PubMed
-
- Bell C.C., Grimm R.J. (1969) Discharge properties of Purkinje cells recorded on single and double microelectrodes. J. Neurophysiol., 32, 1044–1055. - PubMed
-
- Thach W.T. (1968) Discharge of Purkinje and cerebellar nuclear neurons during rapidly alternating arm movements in the monkey. J. Neurophysiol., 31, 785–797. - PubMed
-
- Thach W.T. (1978) Correlation of neural discharge with pattern and force of muscular activity, joint position, and direction of intended next movement in motor cortex and cerebellum. J. Neurophysiol., 41, 654–676. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
