

A review of the basics of mitochondrial bioenergetics, metabolism, and related signaling pathways in cancer cells: Therapeutic targeting of tumor mitochondria with lipophilic cationic compounds
- PMID: 29017115
- PMCID: PMC5633086
- DOI: 10.1016/j.redox.2017.09.020
A review of the basics of mitochondrial bioenergetics, metabolism, and related signaling pathways in cancer cells: Therapeutic targeting of tumor mitochondria with lipophilic cationic compounds
Erratum in
-
Corrigendum to 'A review of the basics of mitochondrial bioenergetics, metabolism, and related signaling pathways in cancer cells: Therapeutic targeting of tumor mitochondria with lipophilic cationic compounds' [REDOX 14C (2017) 316-327].Redox Biol. 2018 Jun;16:426-427. doi: 10.1016/j.redox.2018.03.001. Epub 2018 Mar 13. Redox Biol. 2018. PMID: 29548725 Free PMC article. No abstract available.
Abstract
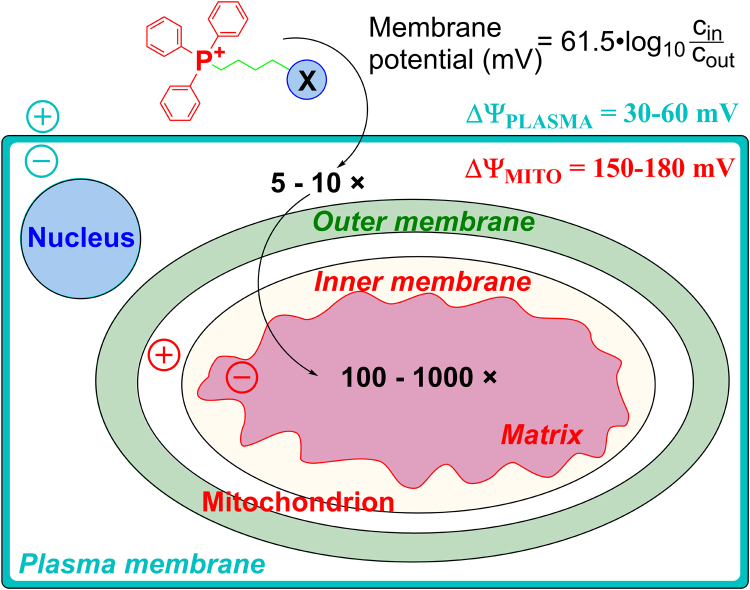
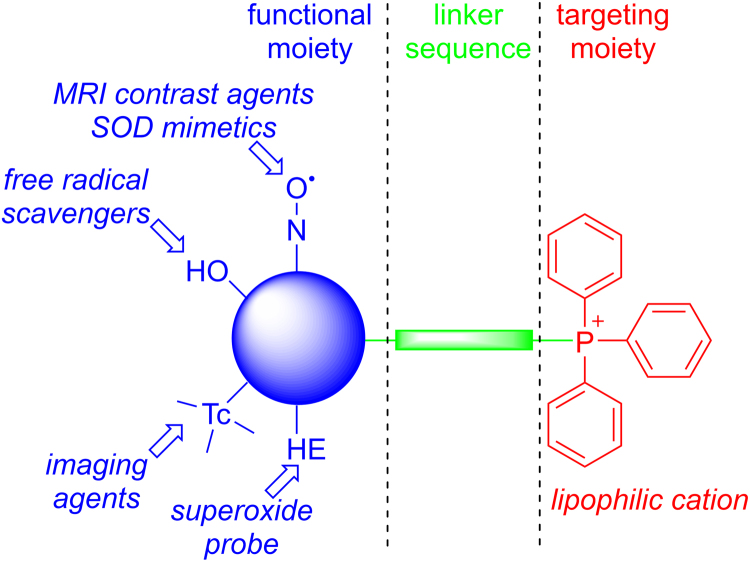
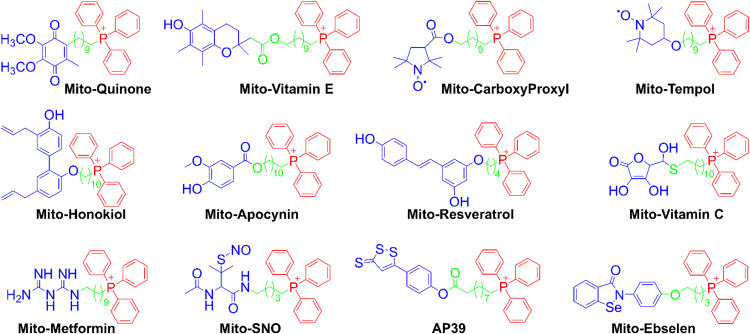
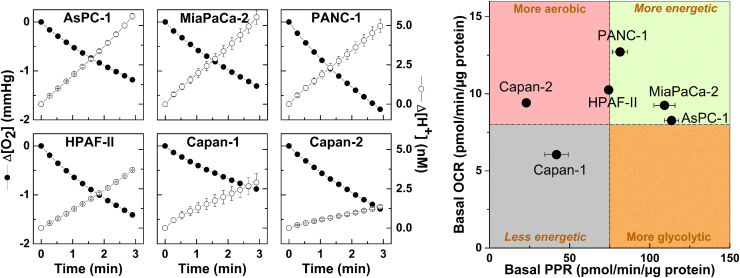
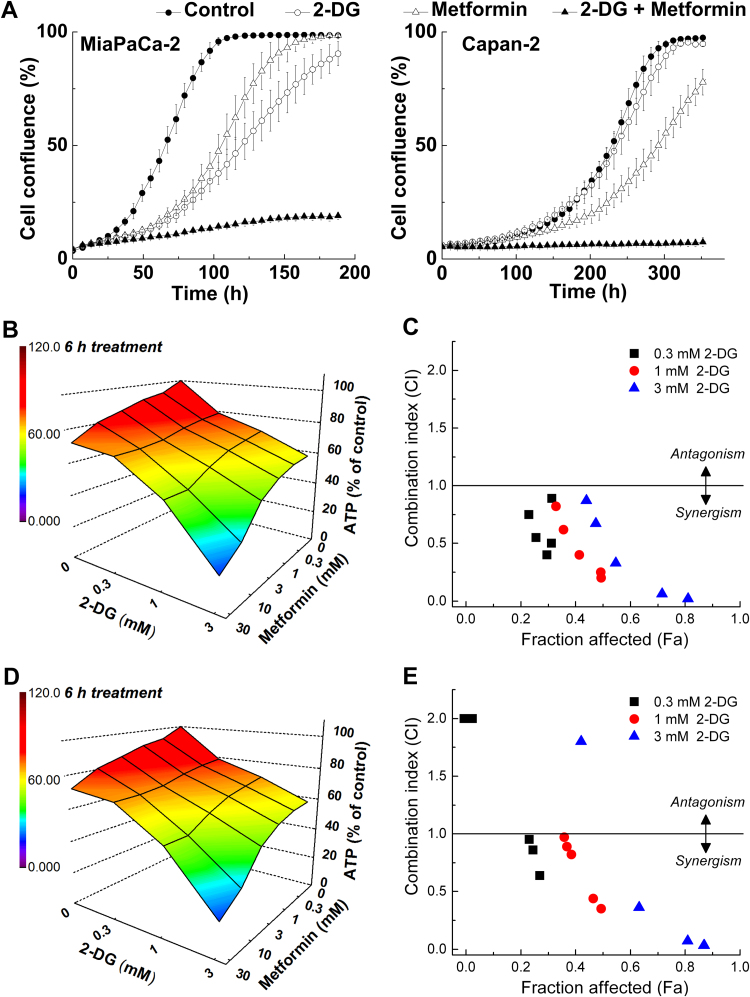
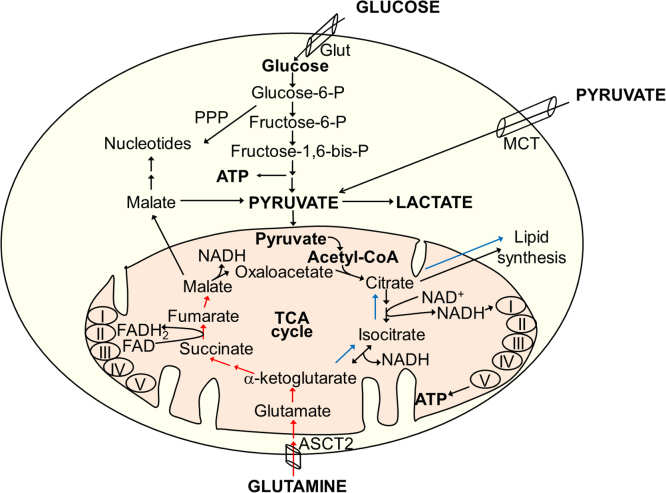
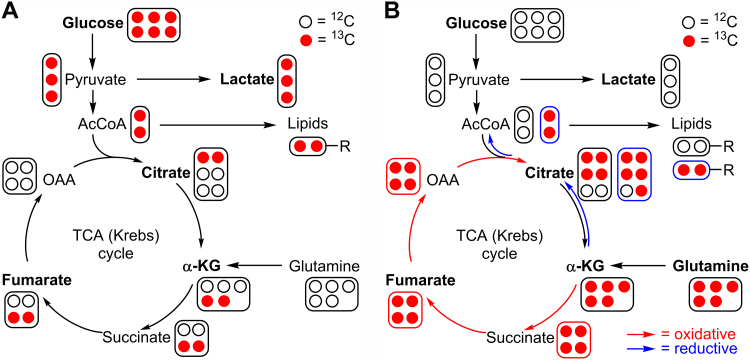
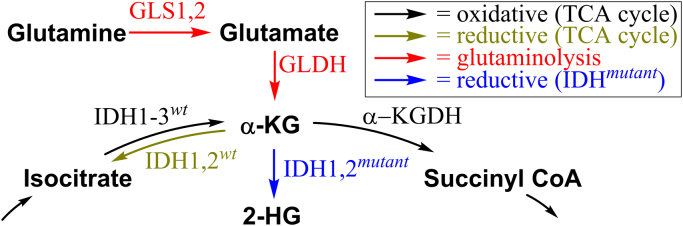
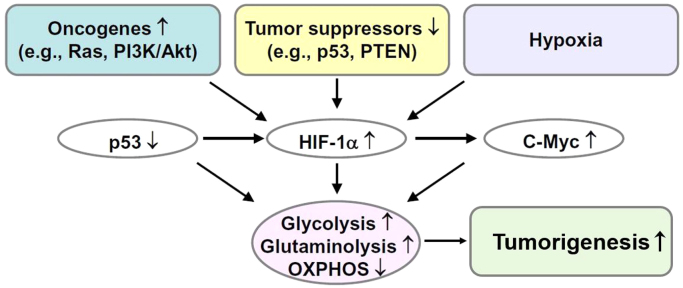
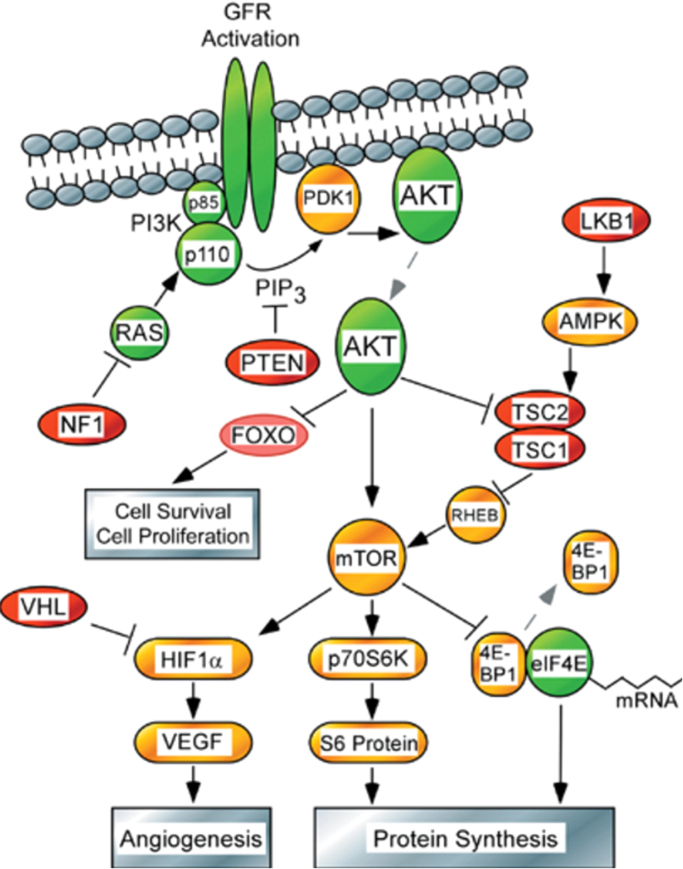
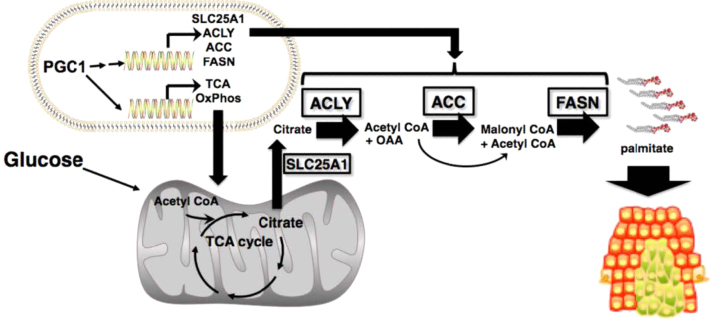
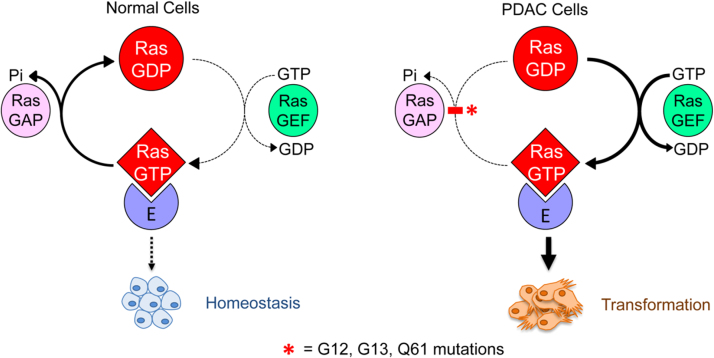
The present review is a sequel to the previous review on cancer metabolism published in this journal. This review focuses on the selective antiproliferative and cytotoxic effects of mitochondria-targeted therapeutics (MTTs) in cancer cells. Emerging research reveals a key role of mitochondrial respiration on tumor proliferation. Previously, a mitochondria-targeted nitroxide was shown to selectively inhibit colon cancer cell proliferation at submicromolar levels. This review is centered on the therapeutic use of MTTs and their bioenergetic profiling in cancer cells. Triphenylphosphonium cation conjugated to a parent molecule (e.g., vitamin-E or chromanol, ubiquinone, and metformin) via a linker alkyl chain is considered an MTT. MTTs selectively and potently inhibit proliferation of cancer cells and, in some cases, induce cytotoxicity. MTTs inhibit mitochondrial complex I activity and induce mitochondrial stress in cancer cells through generation of reactive oxygen species. MTTs in combination with glycolytic inhibitors synergistically inhibit tumor cell proliferation. This review discusses how signaling molecules traditionally linked to tumor cell proliferation affect tumor metabolism and bioenergetics (glycolysis, TCA cycle, and glutaminolysis).
Keywords: Coenzyme Q(10); Extracellular acidification rate; Oxygen consumption rate; Pancreatic ductal adenocarcinoma; Triphenylphosphonium cation.
Copyright © 2017 The Authors. Published by Elsevier B.V. All rights reserved.
Figures
References
-
- Hsu P.P., Sabatini D.M. Cancer cell metabolism: Warburg and beyond. Cell. 2008;134(5):703–707. - PubMed
-
- Warburg O. On the origin of cancer cells. Science. 1956;123(3191):309–314. - PubMed
-
- Beckner M.E., Gobbel G.T., Abounader R., Burovic F., Agostino N.R., Laterra J., Pollack I.F. Glycolytic glioma cells with active glycogen synthase are sensitive to PTEN and inhibitors of PI3K and gluconeogenesis. Lab. Invest. 2005;85(12):1457–1470. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
