

How activating mutations affect MEK1 regulation and function
- PMID: 29018093
- PMCID: PMC5704466
- DOI: 10.1074/jbc.C117.806067
How activating mutations affect MEK1 regulation and function
Abstract
The MEK1 kinase directly phosphorylates ERK2, after the activation loop of MEK1 is itself phosphorylated by Raf. Studies over the past decade have revealed a large number of disease-related mutations in the MEK1 gene that lead to tumorigenesis and abnormal development. Several of these mutations result in MEK1 constitutive activity, but how they affect MEK1 regulation and function remains largely unknown. Here, we address these questions focusing on two pathogenic variants of the Phe-53 residue, which maps to the well-characterized negative regulatory region of MEK1. We found that these variants are phosphorylated by Raf faster than the wild-type enzyme, and this phosphorylation further increases their enzymatic activity. However, the maximal activities of fully phosphorylated wild-type and mutant enzymes are indistinguishable. On the basis of available structural information, we propose that the activating substitutions destabilize the inactive conformation of MEK1, resulting in its constitutive activity and making it more prone to Raf-mediated phosphorylation. Experiments in zebrafish revealed that the effects of activating variants on embryonic development reflect the joint control of the negative regulatory region and activating phosphorylation. Our results underscore the complexity of the effects of activating mutations on signaling systems, even at the level of a single protein.
Keywords: MEK1; Raf kinase; cancer biology; mitogen-activated protein kinase (MAPK); protein phosphorylation; zebrafish.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
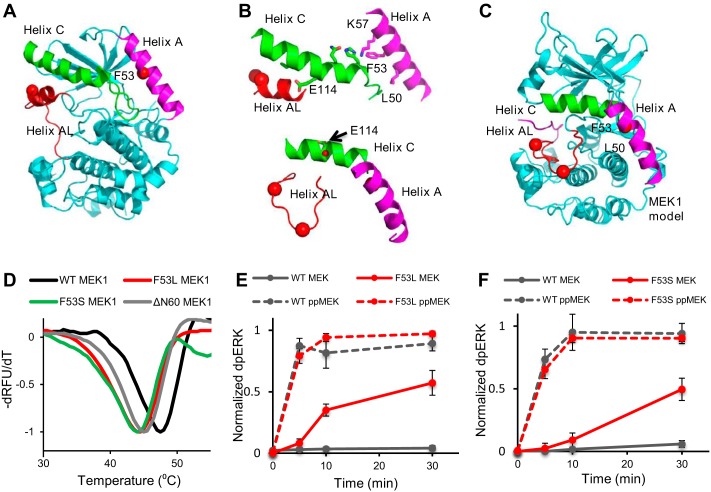
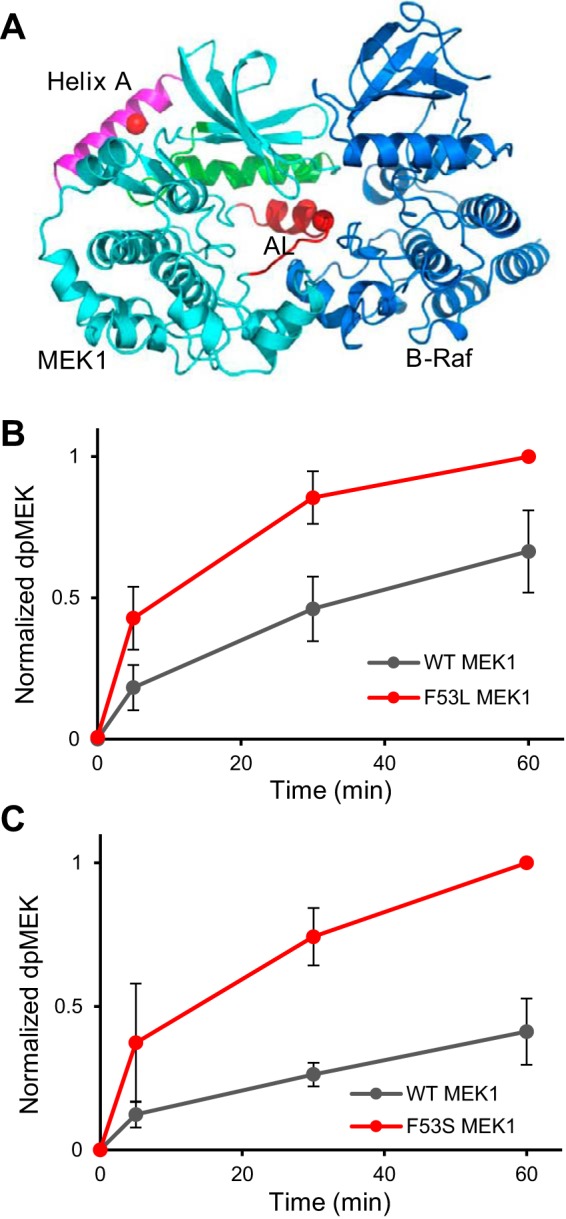
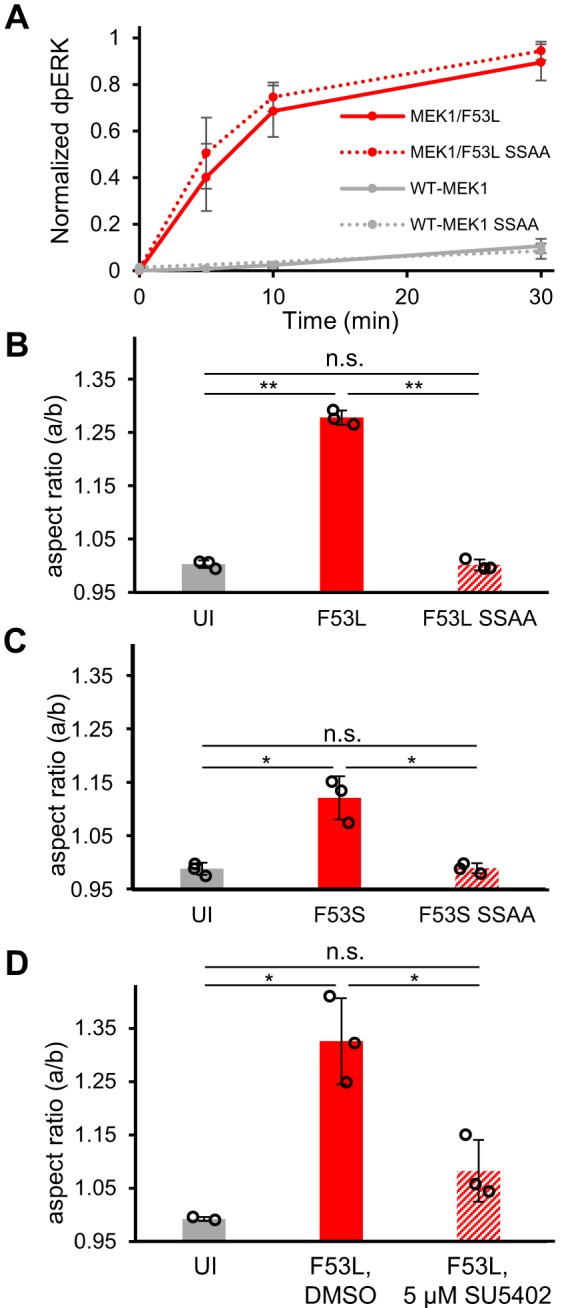
References
-
- Seger R., Ahn N. G., Posada J., Munar E. S., Jensen A. M., Cooper J. A., Cobb M. H., and Krebs E. G. (1992) Purification and characterization of mitogen-activated protein kinase. Activator(s) from epidermal growth factor-stimulated A431 cells. J. Biol. Chem. 267, 14373–14381 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
