

Deconstructing a multiple antibiotic resistance regulation through the quantification of its input function
- PMID: 29018569
- PMCID: PMC5630622
- DOI: 10.1038/s41540-017-0031-2
Deconstructing a multiple antibiotic resistance regulation through the quantification of its input function
Abstract
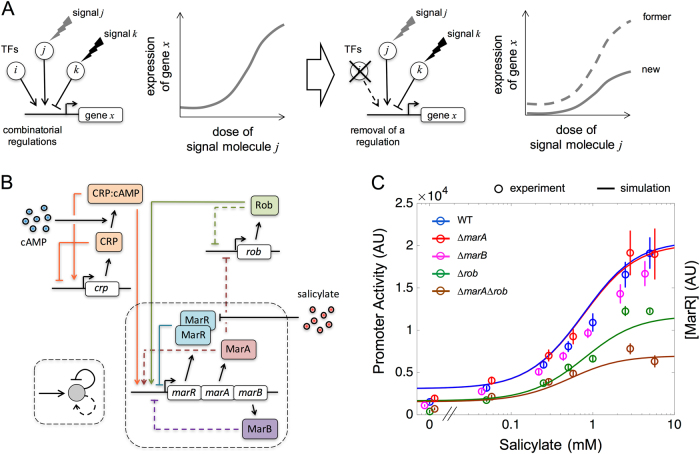
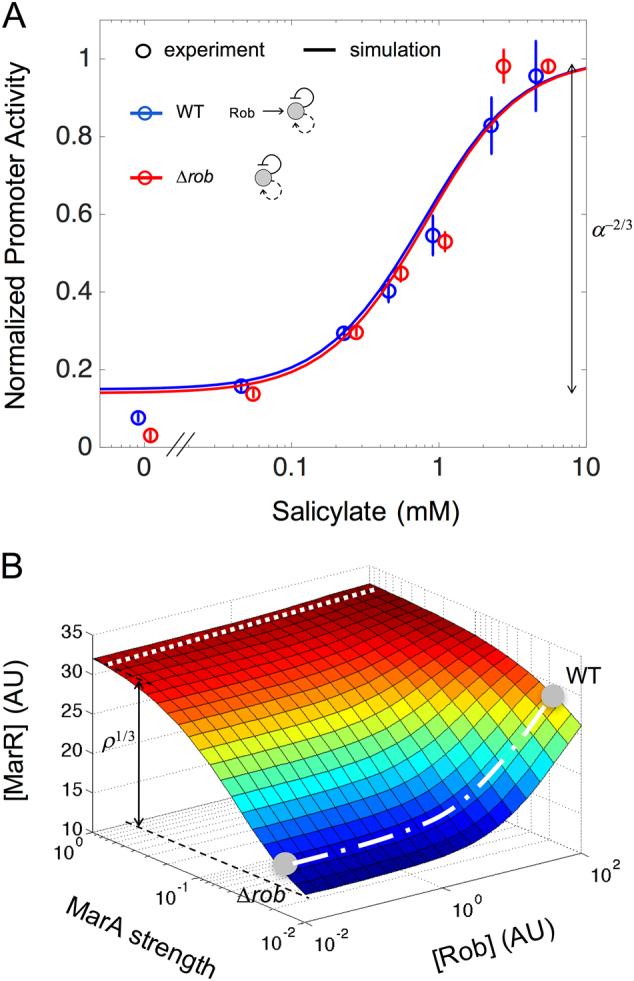
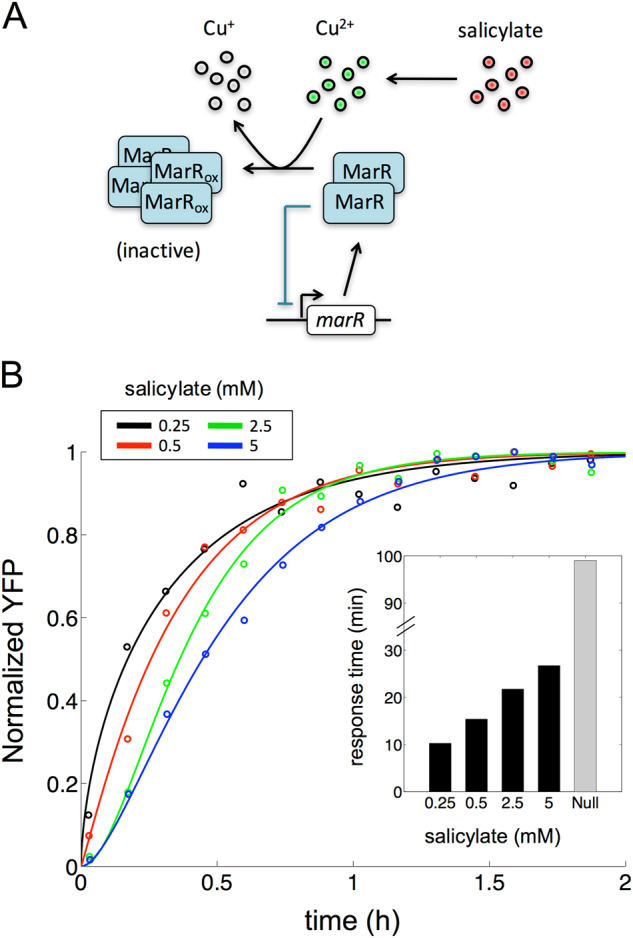
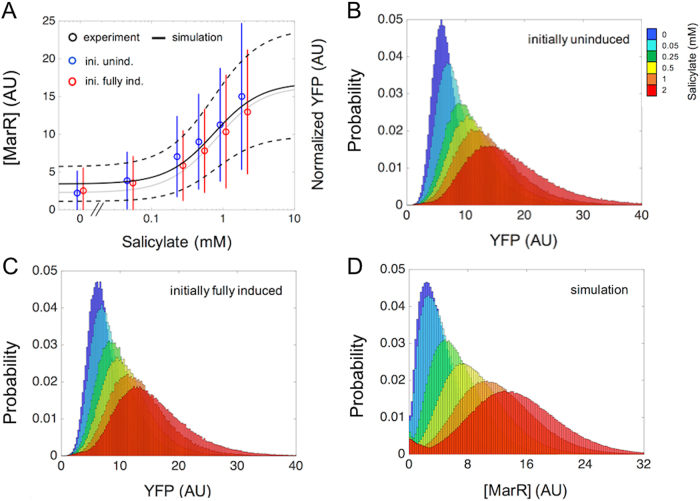
Many essential bacterial responses present complex transcriptional regulation of gene expression. To what extent can the study of these responses substantiate the logic of their regulation? Here, we show how the input function of the genes constituting the response, i.e., the information of how their transcription rates change as function of the signals acting on the regulators, can serve as a quantitative tool to deconstruct the corresponding regulatory logic. To demonstrate this approach, we consider the multiple antibiotic resistance (mar) response in Escherichia coli. By characterizing the input function of its representative genes in wild-type and mutant bacteria, we recognize a dual autoregulation motif as main determinant of the response, which is further adjusted by the interplay with other regulators. We show that basic attributes, like its reaction to a wide range of stress or its moderate expression change, are associated with a strong negative autoregulation, while others, like the buffering of metabolic signals or the lack of memory to previous stress, are related to a weak positive autoregulation. With a mathematical model of the input functions, we identify some constraints fixing the molecular attributes of the regulators, and also notice the relevance of the bicystronic architecture harboring the dual autoregulation that is unique in E. coli. The input function emerges then as a tool to disentangle the rationale behind most of the attributes defining the mar phenotype. Overall, the present study supports the value of characterizing input functions to deconstruct the complexity of regulatory architectures in prokaryotic and eukaryotic systems.
Conflict of interest statement
The authors declare no competing financial interests.
Figures


References
-
- Ptashne, M. & Gann, A. Genes and signals (Cold Spring Harbor Laboratory Press, 2002).
-
- Ptashne, M. Genetic Switch (Cold Spring Harbor Laboratory Press, 2004).
LinkOut - more resources
Full Text Sources
Other Literature Sources
