

Emerging Role of One-Carbon Metabolism and DNA Methylation Enrichment on δ-Containing GABAA Receptor Expression in the Cerebellum of Subjects with Alcohol Use Disorders (AUD)
- PMID: 29020412
- PMCID: PMC5716183
- DOI: 10.1093/ijnp/pyx075
Emerging Role of One-Carbon Metabolism and DNA Methylation Enrichment on δ-Containing GABAA Receptor Expression in the Cerebellum of Subjects with Alcohol Use Disorders (AUD)
Abstract
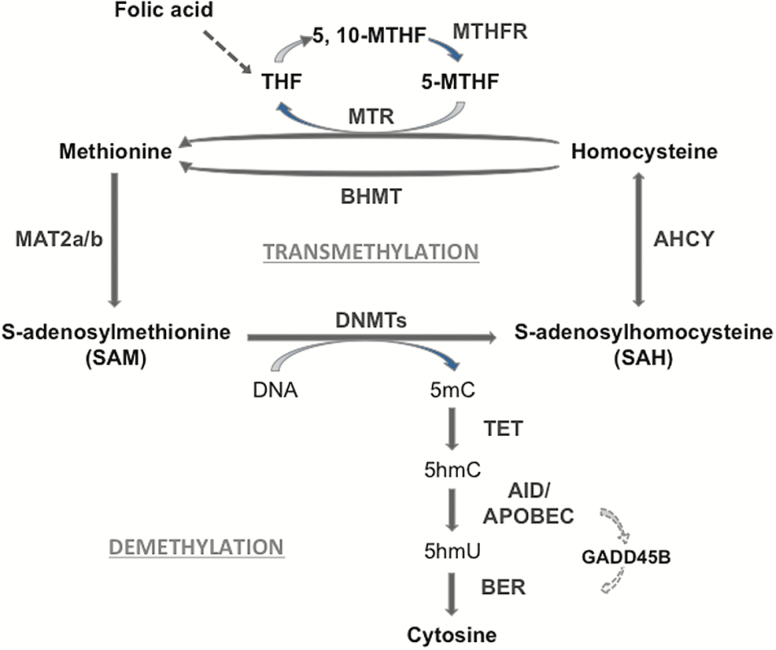
Background: Cerebellum is an area of the brain particularly sensitive to the effects of acute and chronic alcohol consumption. Alcohol exposure decreases cerebellar Purkinje cell output by increasing GABA release from Golgi cells onto extrasynaptic α6/δ-containing GABAA receptors located on glutamatergic granule cells. Here, we studied whether chronic alcohol consumption induces changes in GABAA receptor subunit expression and whether these changes are associated with alterations in epigenetic mechanisms via DNA methylation.
Methods: We used a cohort of postmortem cerebellum from control and chronic alcoholics, here defined as alcohol use disorders subjects (n=25/group). S-adenosyl-methionine/S-adenosyl-homocysteine were measured by high-performance liquid chromatography. mRNA levels of various genes were assessed by reverse transcriptase-quantitative polymerase chain reaction. Promoter methylation enrichment was assessed using methylated DNA immunoprecipitation and hydroxy-methylated DNA immunoprecipitation assays.
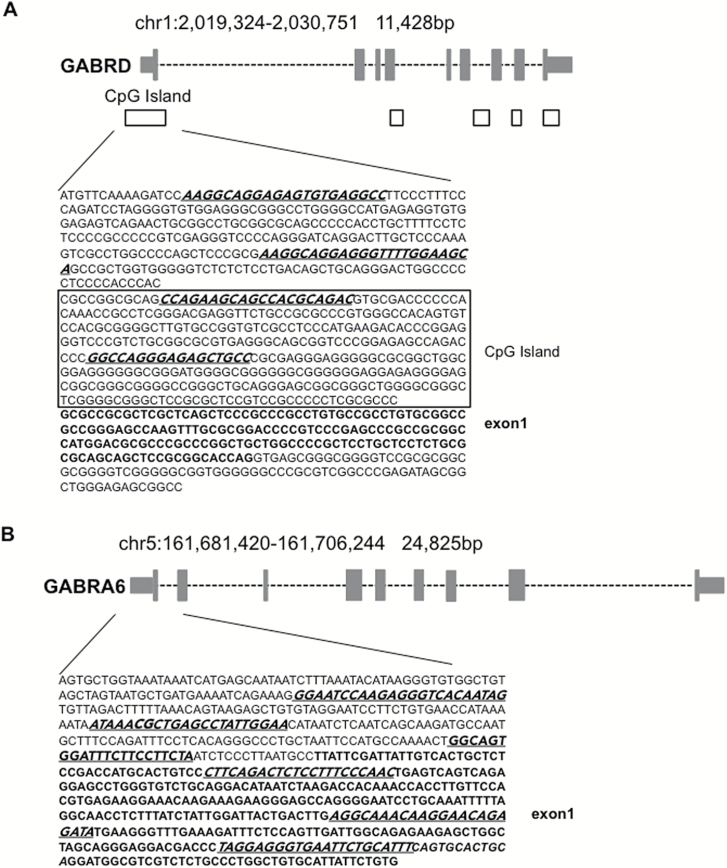
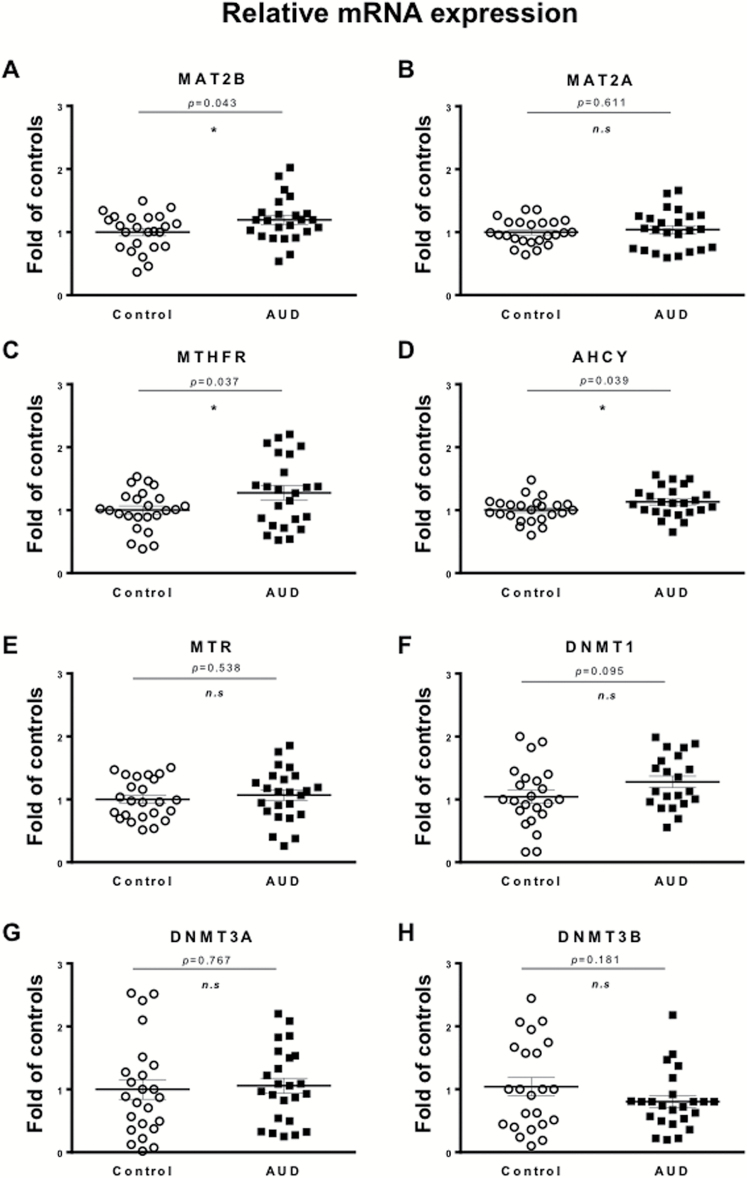
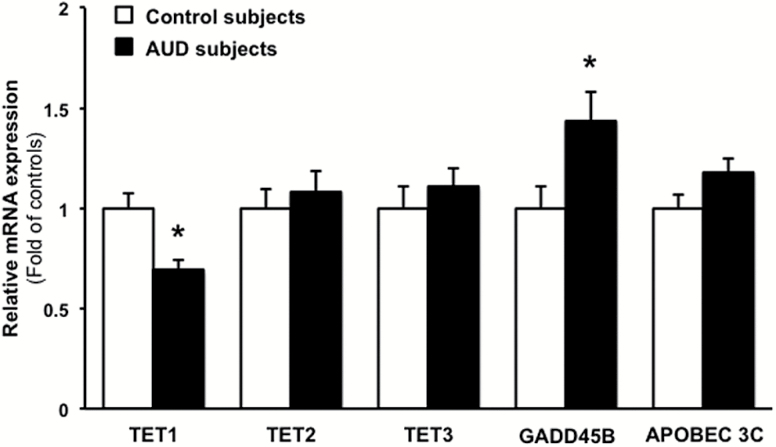
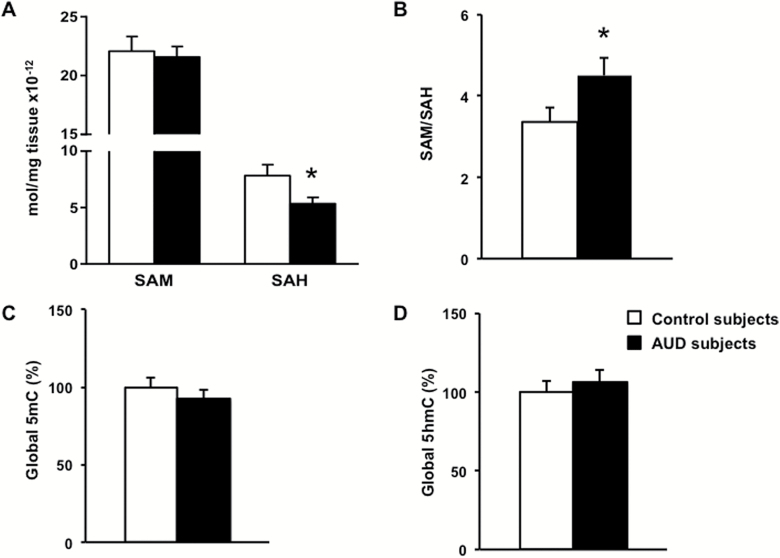
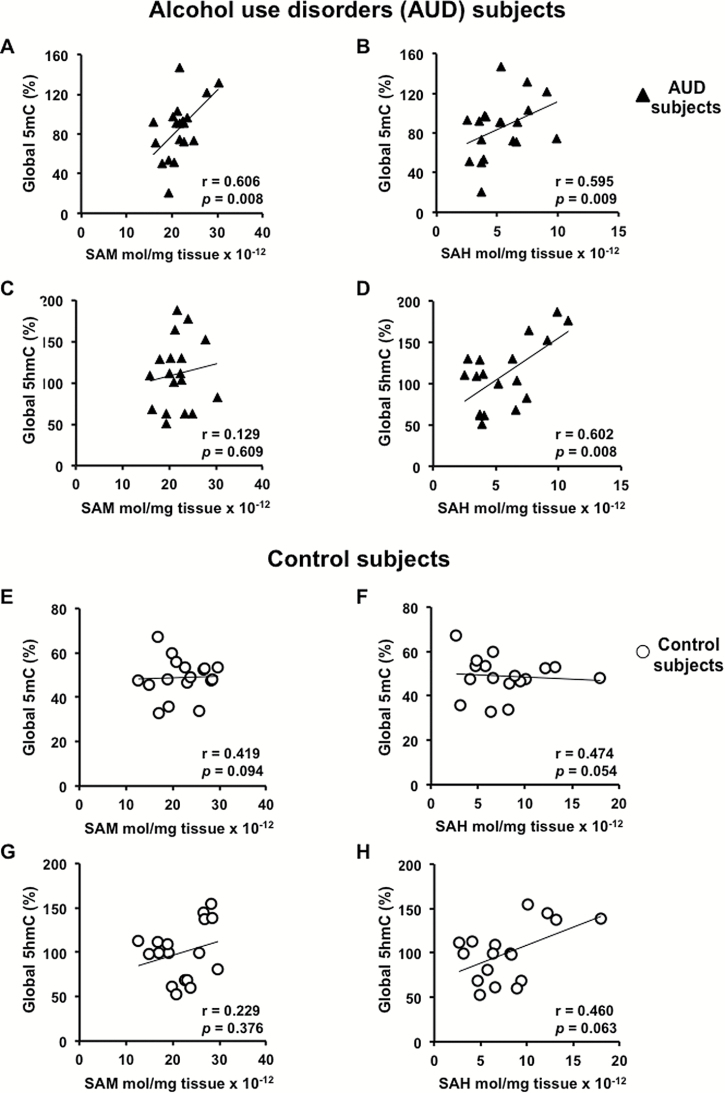
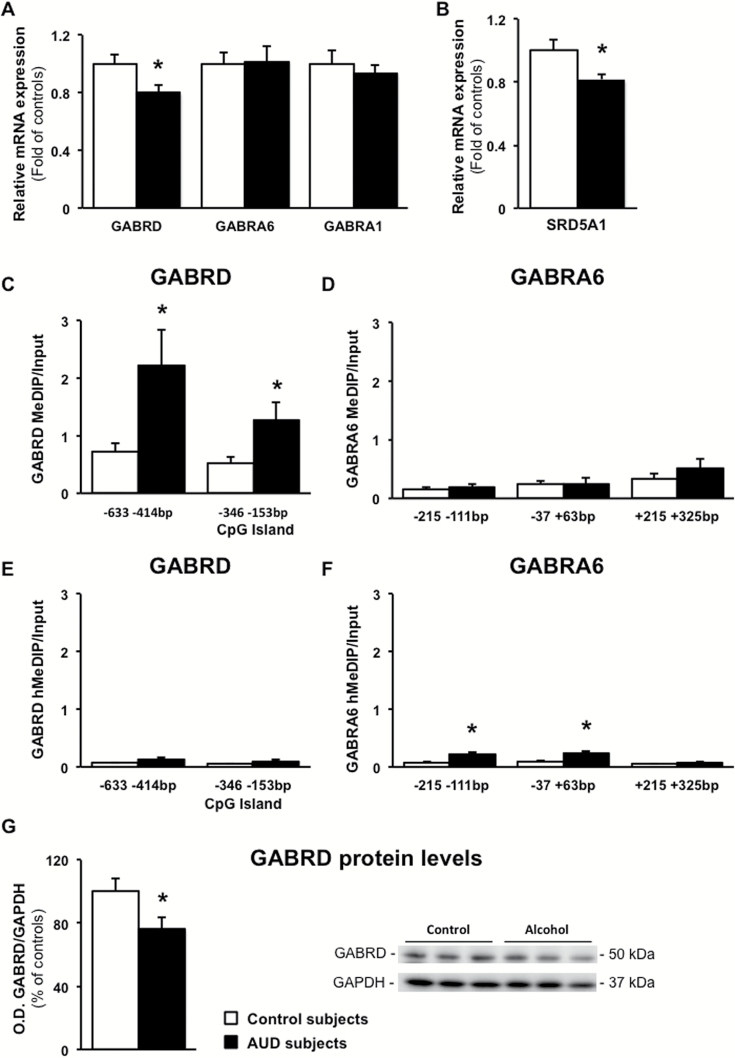
Results: mRNAs encoding key enzymes of 1-carbon metabolism that determine the S-adenosyl-methionine/S-adenosyl-homocysteine ratio were increased, indicating higher "methylation index" in alcohol use disorder subjects. We found that increased methylation of the promoter of the δ subunit GABAA receptor was associated with reduced mRNA and protein levels in the cerebellum of alcohol use disorder subjects. No changes were observed in α1- or α6-containing GABAA receptor subunits. The expression of DNA-methyltransferases (1, 3A, and 3B) was unaltered, whereas the mRNA level of TET1, which participates in the DNA demethylation pathway, was decreased. Hence, increased methylation of the δ subunit GABAA receptor promoter may result from alcohol-induced reduction of DNA demethylation.
Conclusion: Together, these results support the hypothesis that aberrant DNA methylation pathways may be involved in cerebellar pathophysiology of alcoholism. Furthermore, this work provides novel evidence for a central role of DNA methylation mechanisms in the alcohol-induced neuroadaptive changes of human cerebellar GABAA receptor function.
Keywords: GABAA receptor; alcohol; cerebellum; methylation; one-carbon metabolism.
© The Author 2017. Published by Oxford University Press on behalf of CINP.
Figures
Similar articles
-
Chronic Alcohol Exposure Differentially Alters One-Carbon Metabolism in Rat Liver and Brain.Alcohol Clin Exp Res. 2017 Jun;41(6):1105-1111. doi: 10.1111/acer.13382. Epub 2017 Apr 25. Alcohol Clin Exp Res. 2017. PMID: 28369960 Free PMC article.
-
Alterations in the expression of GABAA receptor subunits in cerebellar granule cells after the disruption of the alpha6 subunit gene.Eur J Neurosci. 1999 May;11(5):1685-97. doi: 10.1046/j.1460-9568.1999.00581.x. Eur J Neurosci. 1999. PMID: 10215922
-
Altered gabaa receptor subunit and splice variant expression in rats treated with chronic intermittent ethanol.Alcohol Clin Exp Res. 2001 Jun;25(6):819-28. Alcohol Clin Exp Res. 2001. PMID: 11410716
-
α6-Containing GABAA Receptors: Functional Roles and Therapeutic Potentials.Pharmacol Rev. 2022 Jan;74(1):238-270. doi: 10.1124/pharmrev.121.000293. Pharmacol Rev. 2022. PMID: 35017178 Review.
-
Alcohol: effects on GABAA receptor function and gene expression.Alcohol Alcohol Suppl. 1993;2:331-5. Alcohol Alcohol Suppl. 1993. PMID: 7748320 Review.
Cited by
-
Dual methylation and hydroxymethylation study of alcohol use disorder.Addict Biol. 2022 Mar;27(2):e13114. doi: 10.1111/adb.13114. Epub 2021 Nov 17. Addict Biol. 2022. PMID: 34791764 Free PMC article.
-
Epigenetic drugs and psychedelics as emerging therapies for alcohol use disorder: insights from preclinical studies.J Neural Transm (Vienna). 2024 May;131(5):525-561. doi: 10.1007/s00702-024-02757-3. Epub 2024 Mar 30. J Neural Transm (Vienna). 2024. PMID: 38554193 Review.
-
Advances in DNA, histone, and RNA methylation mechanisms in the pathophysiology of alcohol use disorder.Adv Drug Alcohol Res. 2023 Feb 15;3:10871. doi: 10.3389/adar.2023.10871. eCollection 2023. Adv Drug Alcohol Res. 2023. PMID: 38389820 Free PMC article. Review.
-
Data-driven study on resting-state functional magnetic resonance imaging during early abstinence of alcohol dependence in male patients and its predictive value for relapse.BMC Psychiatry. 2022 Feb 22;22(1):143. doi: 10.1186/s12888-022-03782-w. BMC Psychiatry. 2022. PMID: 35193538 Free PMC article.
-
Is DNA methylation in the brain a mechanism of alcohol use disorder?Front Behav Neurosci. 2023 Jan 26;17:957203. doi: 10.3389/fnbeh.2023.957203. eCollection 2023. Front Behav Neurosci. 2023. PMID: 36778133 Free PMC article. Review.
References
-
- American Psychiatric Association (1994) Diagnostic and Statistical Manual of Mental Disorders, 4th ed Washington, DC: American Psychiatric Association.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
