

Stochastic Community Assembly: Does It Matter in Microbial Ecology?
- PMID: 29021219
- PMCID: PMC5706748
- DOI: 10.1128/MMBR.00002-17
Stochastic Community Assembly: Does It Matter in Microbial Ecology?
Abstract
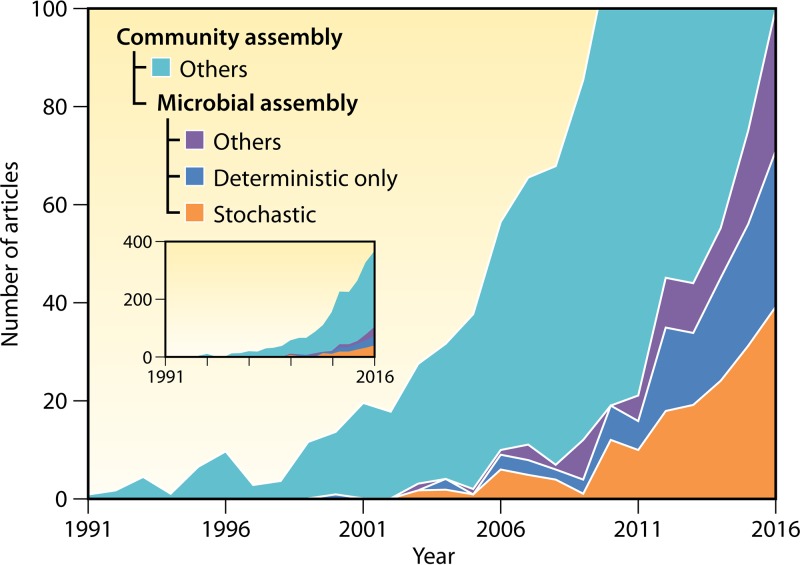
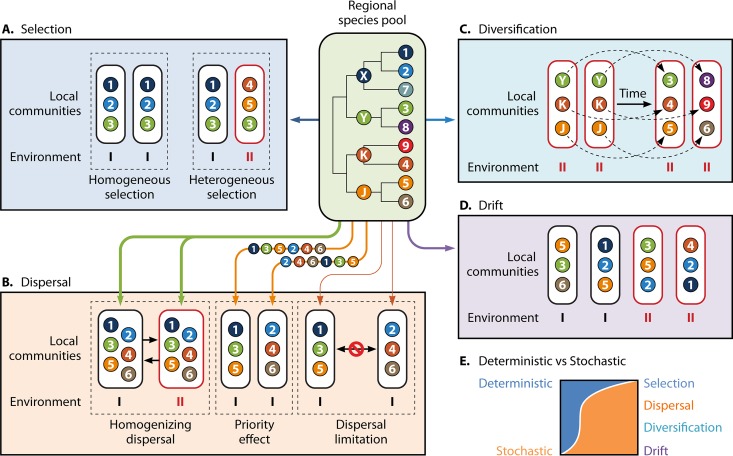
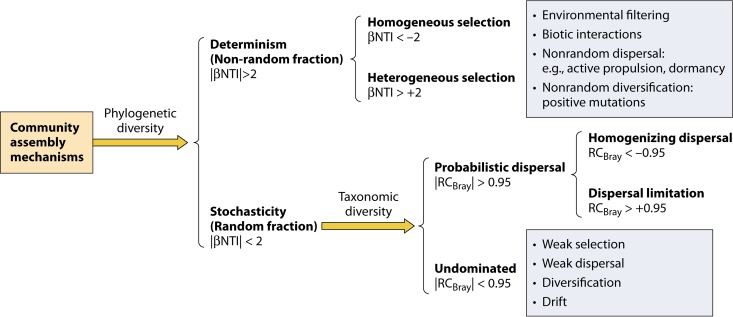
Understanding the mechanisms controlling community diversity, functions, succession, and biogeography is a central, but poorly understood, topic in ecology, particularly in microbial ecology. Although stochastic processes are believed to play nonnegligible roles in shaping community structure, their importance relative to deterministic processes is hotly debated. The importance of ecological stochasticity in shaping microbial community structure is far less appreciated. Some of the main reasons for such heavy debates are the difficulty in defining stochasticity and the diverse methods used for delineating stochasticity. Here, we provide a critical review and synthesis of data from the most recent studies on stochastic community assembly in microbial ecology. We then describe both stochastic and deterministic components embedded in various ecological processes, including selection, dispersal, diversification, and drift. We also describe different approaches for inferring stochasticity from observational diversity patterns and highlight experimental approaches for delineating ecological stochasticity in microbial communities. In addition, we highlight research challenges, gaps, and future directions for microbial community assembly research.
Keywords: community assembly; ecological drift; ecological processes; ecological stochasticity; microbial communities.
Copyright © 2017 American Society for Microbiology.
Figures


References
-
- Lyashevska O, Farnsworth KD. 2012. How many dimensions of biodiversity do we need? Ecol Indic 18:485–492. doi: 10.1016/j.ecolind.2011.12.016. - DOI
-
- Cardinale BJ, Duffy JE, Gonzalez A, Hooper DU, Perrings C, Venail P, Narwani A, Mace GM, Tilman D, Wardle DA, Kinzig AP, Daily GC, Loreau M, Grace JB, Larigauderie A, Srivastava DS, Naeem S. 2012. Biodiversity loss and its impact on humanity. Nature 486:59–67. doi: 10.1038/nature11148. - DOI - PubMed
-
- IPCC Core Writing Team, Pachauri RK, Meyer LA (ed). 2014. Climate change 2014: synthesis, report. Contribution of Working Groups I, II and III to the fifth assessment report of the Intergovernmental Panel on Climate Change. IPCC, Geneva, Switzerland.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
