

Two distinct transcription termination modes dictated by promoters
- PMID: 29021241
- PMCID: PMC5695088
- DOI: 10.1101/gad.301093.117
Two distinct transcription termination modes dictated by promoters
Abstract
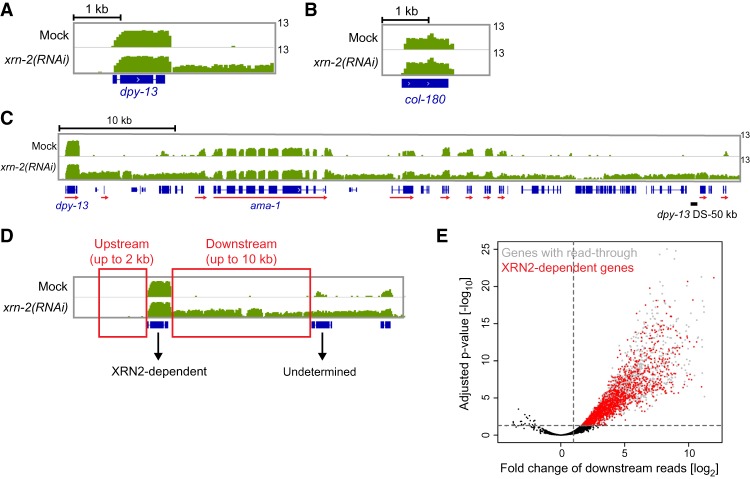
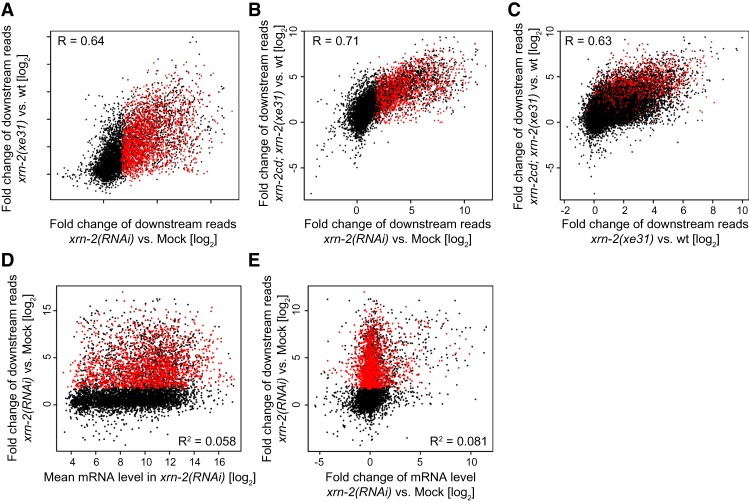
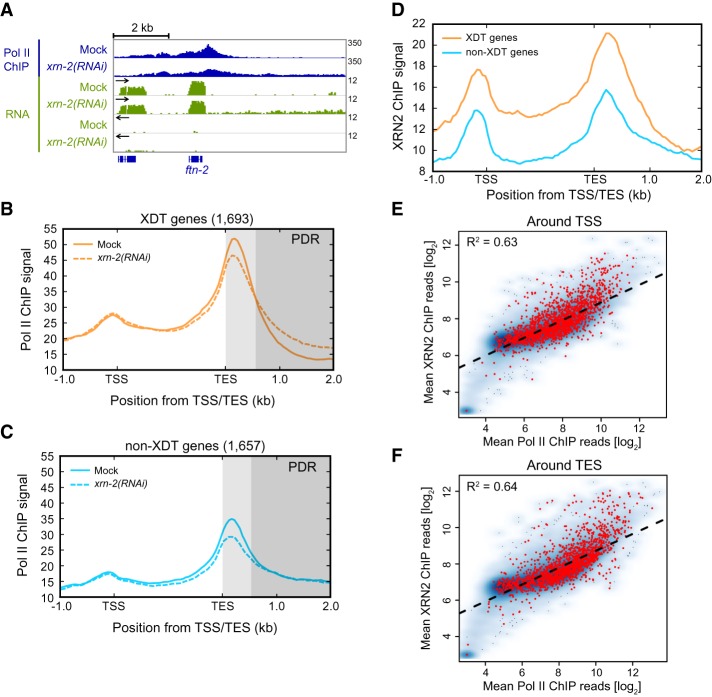
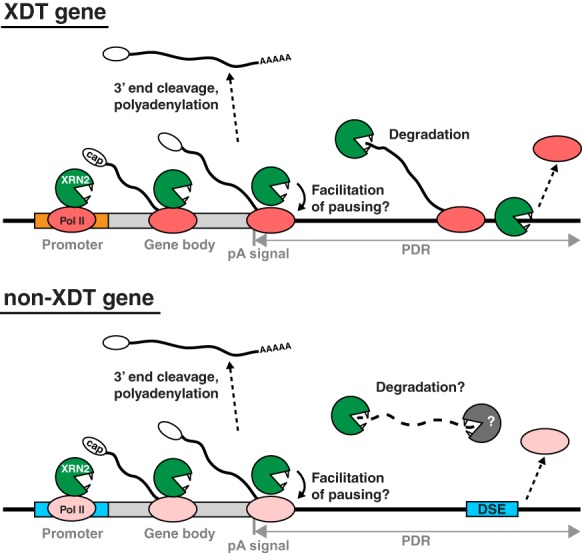
Transcription termination determines the ends of transcriptional units and thereby ensures the integrity of the transcriptome and faithful gene regulation. Studies in yeast and human cells have identified the exoribonuclease XRN2 as a key termination factor for protein-coding genes. Here we performed a genome-wide investigation of RNA polymerase II (Pol II) transcription termination in XRN2-deficient Caenorhabditis elegans and observed two distinct modes of termination. Although a subset of genes requires XRN2, termination of other genes appears both independent of, and refractory to, XRN2. XRN2 independence is not merely a consequence of failure to recruit XRN2, since XRN2 is present on-and promotes Pol II accumulation near the polyadenylation sites of-both gene classes. Unexpectedly, promoters instruct the choice of termination mode, but XRN2-independent termination additionally requires a compatible region downstream from the 3' end cleavage site. Hence, different termination mechanisms may work with different configurations of Pol II complexes dictated by promoters.
Keywords: RNA polymerase II; XRN2, Caenorhabditis elegans; transcription termination.
© 2017 Miki et al.; Published by Cold Spring Harbor Laboratory Press.
Figures

Comment in
-
Transcription: Promoters dictate termination mode.Nat Rev Genet. 2017 Dec;18(12):704. doi: 10.1038/nrg.2017.93. Epub 2017 Oct 30. Nat Rev Genet. 2017. PMID: 29082914 No abstract available.
References
-
- Baejen C, Andreani J, Torkler P, Battaglia S, Schwalb B, Lidschreiber M, Maier KC, Boltendahl A, Rus P, Esslinger S, et al. 2017. Genome-wide analysis of RNA polymerase II termination at protein-coding genes. Mol Cell 66: 38–49. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases