

Involvement of the exomer complex in the polarized transport of Ena1 required for Saccharomyces cerevisiae survival against toxic cations
- PMID: 29021337
- PMCID: PMC5706994
- DOI: 10.1091/mbc.E17-09-0549
Involvement of the exomer complex in the polarized transport of Ena1 required for Saccharomyces cerevisiae survival against toxic cations
Abstract
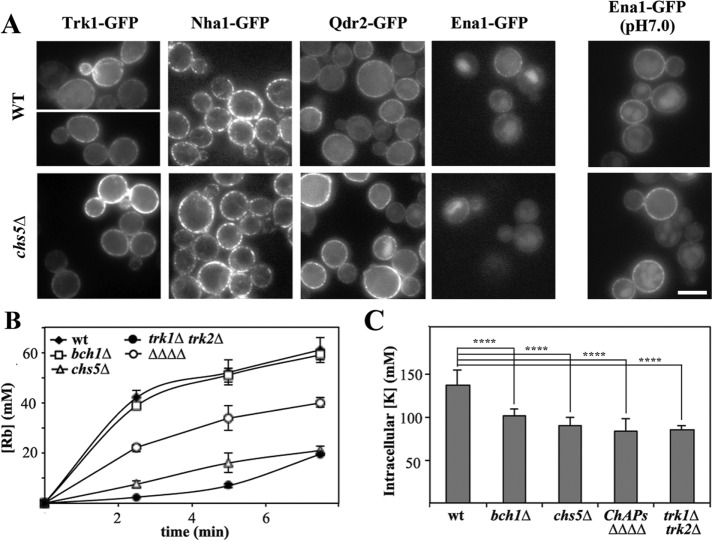
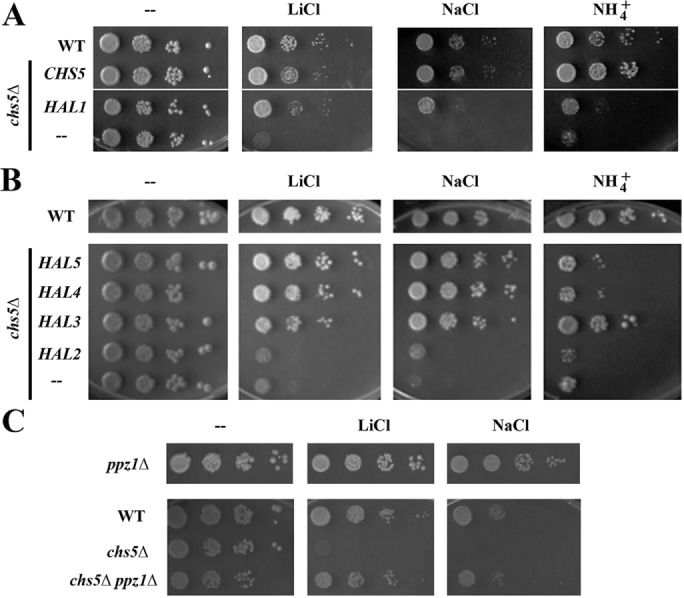
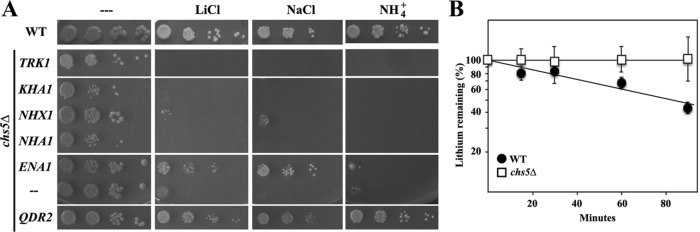
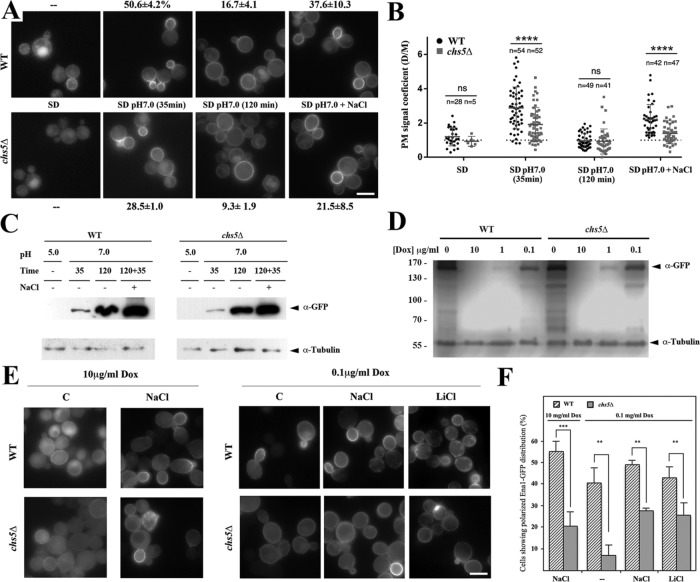
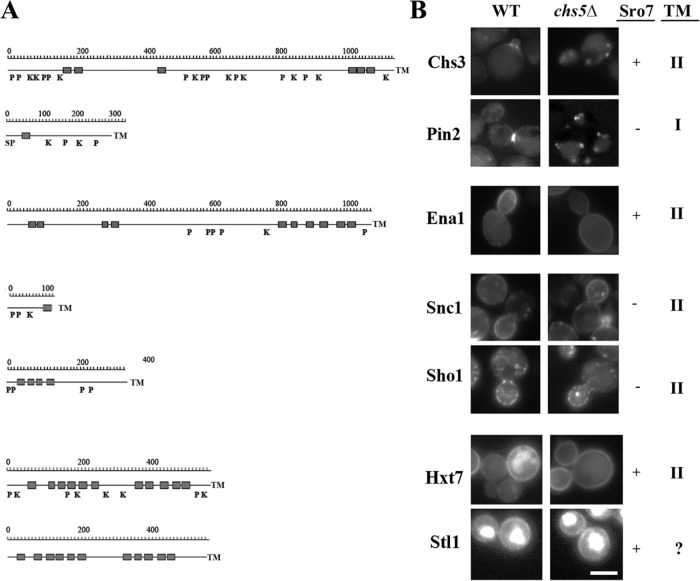
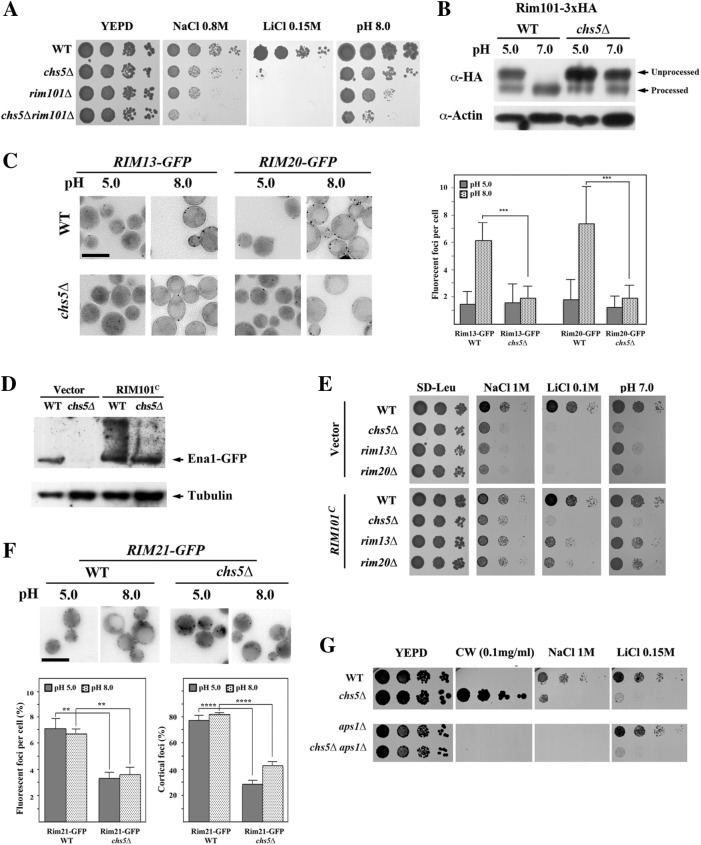
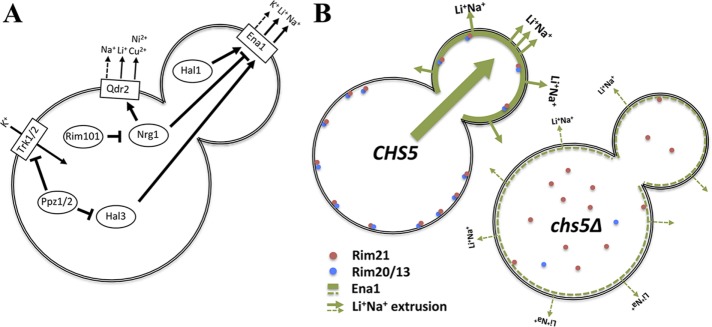
Exomer is an adaptor complex required for the direct transport of a selected number of cargoes from the trans-Golgi network (TGN) to the plasma membrane in Saccharomyces cerevisiae However, exomer mutants are highly sensitive to increased concentrations of alkali metal cations, a situation that remains unexplained by the lack of transport of any known cargoes. Here we identify several HAL genes that act as multicopy suppressors of this sensitivity and are connected to the reduced function of the sodium ATPase Ena1. Furthermore, we find that Ena1 is dependent on exomer function. Even though Ena1 can reach the plasma membrane independently of exomer, polarized delivery of Ena1 to the bud requires functional exomer. Moreover, exomer is required for full induction of Ena1 expression after cationic stress by facilitating the plasma membrane recruitment of the molecular machinery involved in Rim101 processing and activation of the RIM101 pathway in response to stress. Both the defective localization and the reduced levels of Ena1 contribute to the sensitivity of exomer mutants to alkali metal cations. Our work thus expands the spectrum of exomer-dependent proteins and provides a link to a more general role of exomer in TGN organization.
© 2017 Anton et al. This article is distributed by The American Society for Cell Biology under license from the author(s). Two months after publication it is available to the public under an Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).
Figures

References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
