

Disruption of Trim9 function abrogates macrophage motility in vivo
- PMID: 29021367
- PMCID: PMC6608060
- DOI: 10.1189/jlb.1A0816-371R
Disruption of Trim9 function abrogates macrophage motility in vivo
Abstract
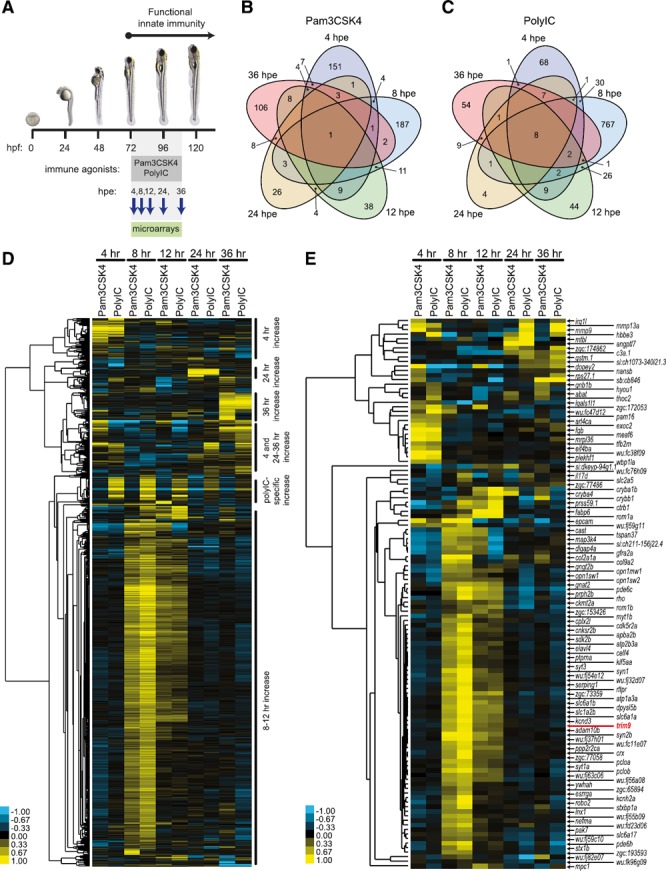
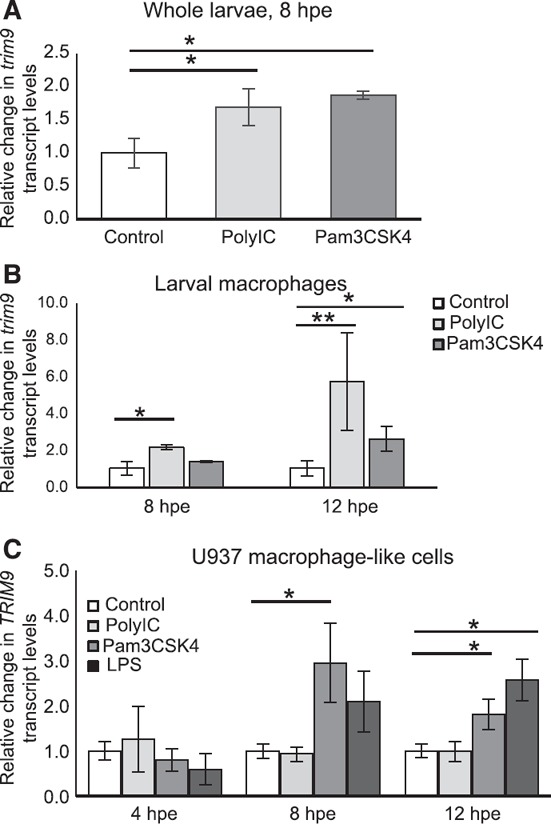
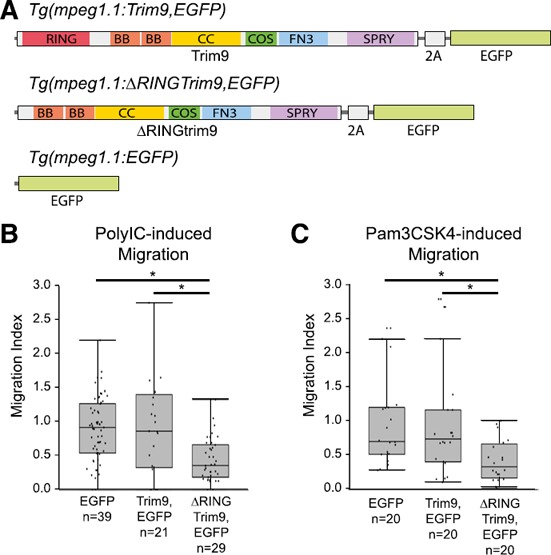
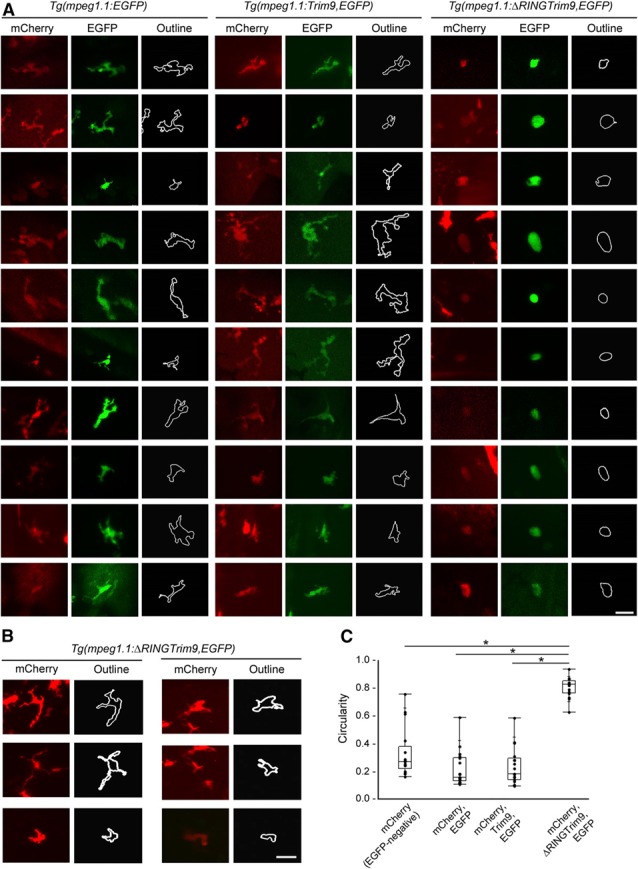
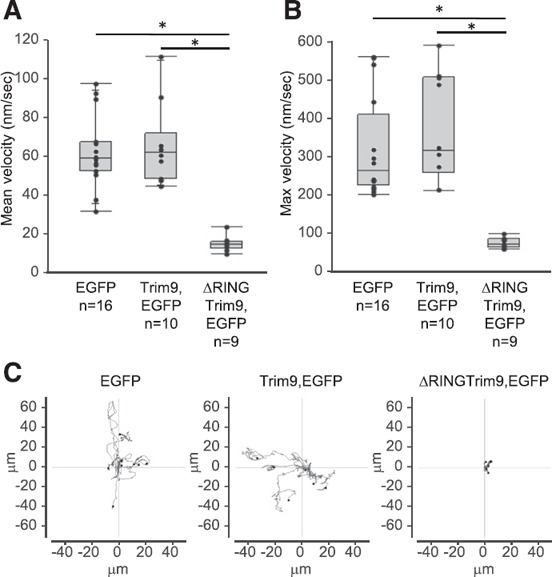
The vertebrate immune response comprises multiple molecular and cellular components that interface to provide defense against pathogens. Because of the dynamic complexity of the immune system and its interdependent innate and adaptive functionality, an understanding of the whole-organism response to pathogen exposure remains unresolved. Zebrafish larvae provide a unique model for overcoming this obstacle, because larvae are protected against pathogens while lacking a functional adaptive immune system during the first few weeks of life. Zebrafish larvae were exposed to immune agonists for various lengths of time, and a microarray transcriptome analysis was executed. This strategy identified known immune response genes, as well as genes with unknown immune function, including the E3 ubiquitin ligase tripartite motif-9 (Trim9). Although trim9 expression was originally described as "brain specific," its expression has been reported in stimulated human Mϕs. In this study, we found elevated levels of trim9 transcripts in vivo in zebrafish Mϕs after immune stimulation. Trim9 has been implicated in axonal migration, and we therefore investigated the impact of Trim9 disruption on Mϕ motility and found that Mϕ chemotaxis and cellular architecture are subsequently impaired in vivo. These results demonstrate that Trim9 mediates cellular movement and migration in Mϕs as well as neurons.
Keywords: chemotaxis; leukocyte; ubiquitin; zebrafish.
© Society for Leukocyte Biology.
Figures
References
-
- Casellas, F. , LÓpez‐Vivancos, J. , Vergara, M. , Malagelada, J. (1999) Impact of inflammatory bowel disease on health‐related quality of life. Dig. Dis. 17, 208–218. - PubMed
-
- Cheung, A. M. , Tansey, C. M. , Tomlinson, G. , Diaz‐Granados, N. , Matté, A. , Barr, A. , Mehta, S. , Mazer, C. D. , Guest, C. B. , Stewart, T. E. , Al‐Saidi, F. , Cooper, A. B. , Cook, D. , Slutsky, A. S. , Herridge, M. S. (2006) Two‐year outcomes, health care use, and costs of survivors of acute respiratory distress syndrome. Am. J. Respir. Crit. Care Med. 174, 538–544. - PubMed
-
- Lieschke, G. J. , Oates, A. C. , Crowhurst, M. O. , Ward, A. C. , Layton, J. E. (2001) Morphologic and functional characterization of granulocytes and macrophages in embryonic and adult zebrafish. Blood 98, 3087–3096. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
