

The cholesterol metabolite 27 hydroxycholesterol facilitates breast cancer metastasis through its actions on immune cells
- PMID: 29021522
- PMCID: PMC5636879
- DOI: 10.1038/s41467-017-00910-z
The cholesterol metabolite 27 hydroxycholesterol facilitates breast cancer metastasis through its actions on immune cells
Abstract
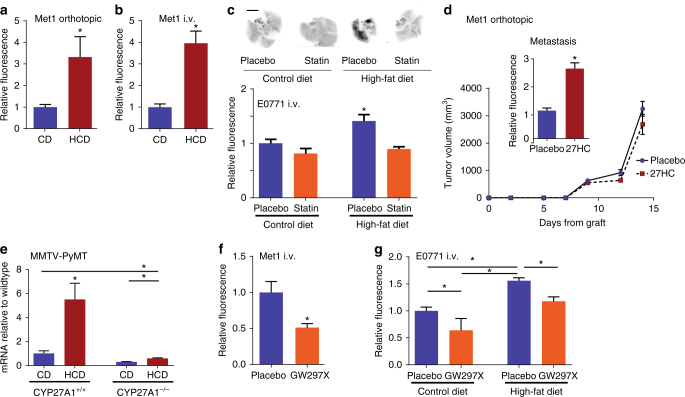
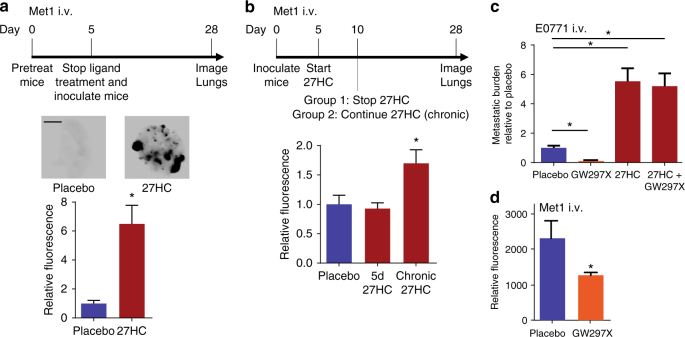
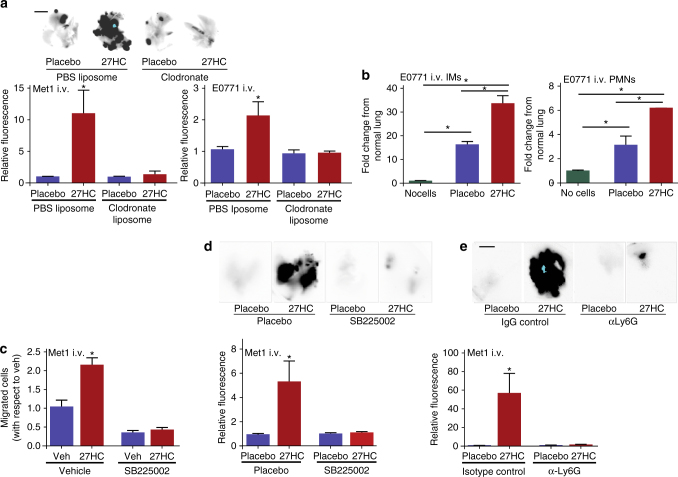
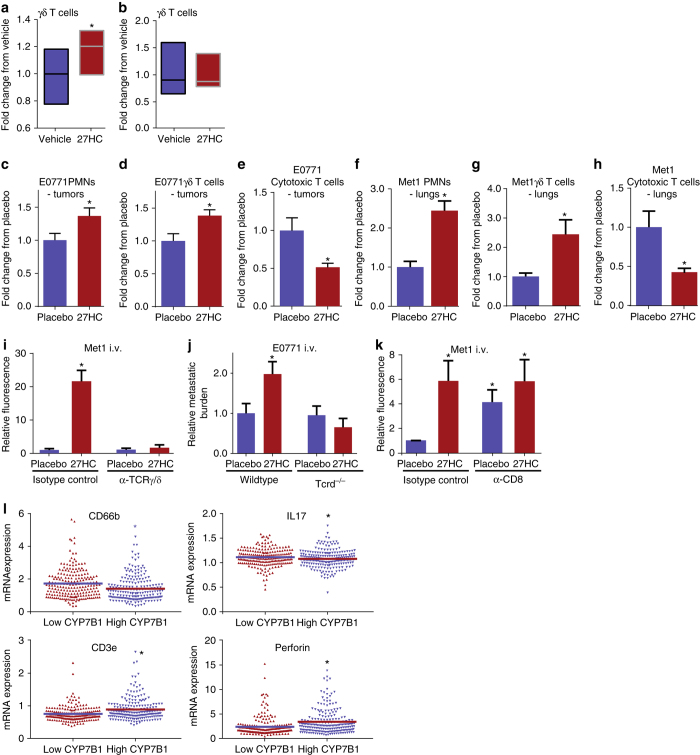
Obesity and elevated circulating cholesterol are risk factors for breast cancer recurrence, while the use of statins, cholesterol biosynthesis inhibitors widely used for treating hypercholesterolemia, is associated with improved disease-free survival. Here, we show that cholesterol mediates the metastatic effects of a high-fat diet via its oxysterol metabolite, 27-hydroxycholesterol. Ablation or inhibition of CYP27A1, the enzyme responsible for the rate-limiting step in 27-hydroxycholesterol biosynthesis, significantly reduces metastasis in relevant animal models of cancer. The robust effects of 27-hydroxycholesterol on metastasis requires myeloid immune cell function, and it was found that this oxysterol increases the number of polymorphonuclear-neutrophils and γδ-T cells at distal metastatic sites. The pro-metastatic actions of 27-hydroxycholesterol requires both polymorphonuclear-neutrophils and γδ-T cells, and 27-hydroxycholesterol treatment results in a decreased number of cytotoxic CD8+T lymphocytes. Therefore, through its actions on γδ-T cells and polymorphonuclear-neutrophils, 27-hydroxycholesterol functions as a biochemical mediator of the metastatic effects of hypercholesterolemia.High cholesterol is a risk factor for breast cancer recurrence. Here the authors show that cholesterol promotes breast cancer metastasis via its metabolite 27-hydroxycholesterol (27HC) that acts on immune myeloid cells residing at the distal metastatic sites, thus promoting an immune suppressive environment.
Conflict of interest statement
E.R.N. and D.P.M. have applied for a patent covering the use of CYP27A1 inhibitors, statins, or LXR antagonists alone or in combination with conventional therapy for the treatment of breast cancer. Title: “Use of cyp27a1 inhibitors, statin, or lxr antagonists alone or in combination with conventional therapy for the treatment of breast cancer”. Inventors: D.P.M. and E.R.N. Applicant: Duke University. Filing Date: Jan. 3, 2014. Application Number: PCT/US2014/010179. The remaining authors declare no competing financial interests.
Figures
References
-
- Centers fo Disease Control and Prevention Prevalence of abnormal lipid levels among youths- United States, 1999–2006. MMWR. 2010;59:29–33. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
