

Potassium in the Grape (Vitis vinifera L.) Berry: Transport and Function
- PMID: 29021796
- PMCID: PMC5623721
- DOI: 10.3389/fpls.2017.01629
Potassium in the Grape (Vitis vinifera L.) Berry: Transport and Function
Abstract
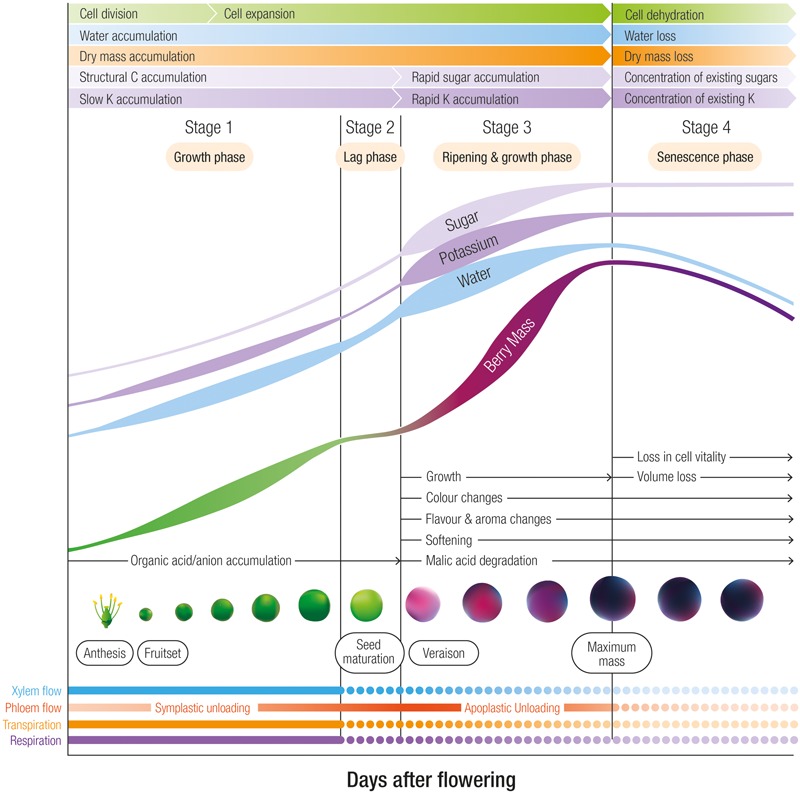
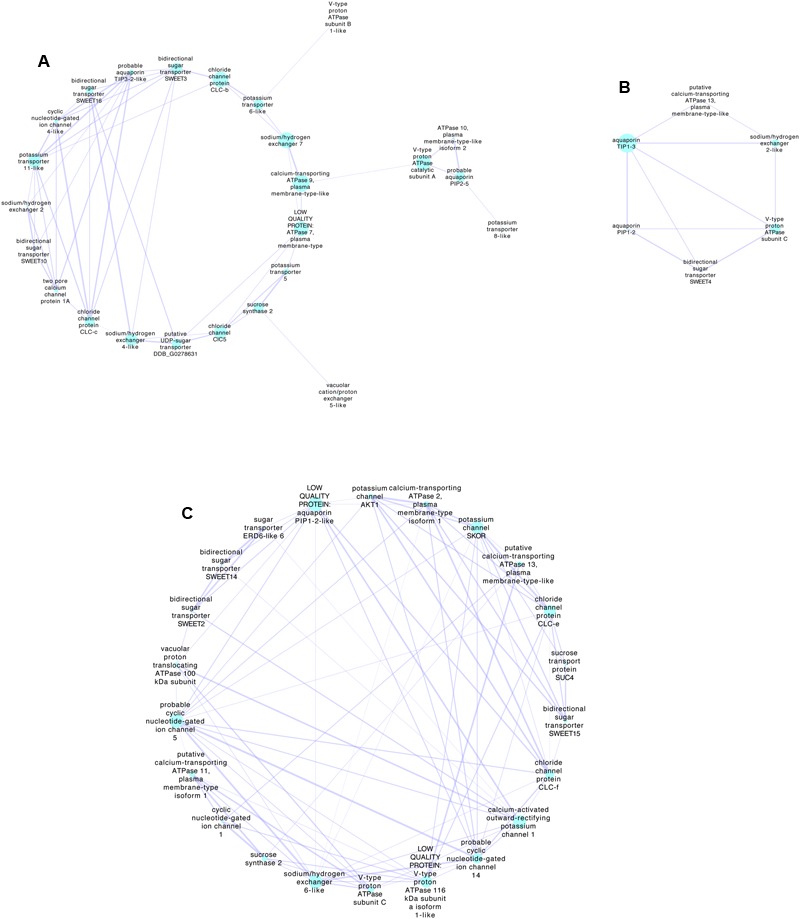
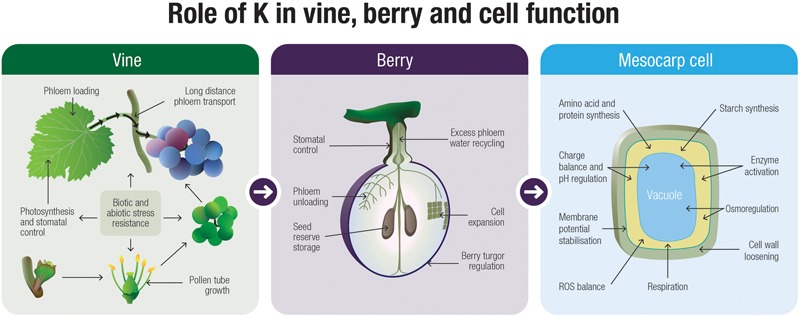
K+ is the most abundant cation in the grape berry. Here we focus on the most recent information in the long distance transport and partitioning of K+ within the grapevine and postulate on the potential role of K+ in berry sugar accumulation, berry water relations, cellular growth, disease resistance, abiotic stress tolerance and mitigating senescence. By integrating information from several different plant systems we have been able to generate new hypotheses on the integral functions of this predominant cation and to improve our understanding of how these functions contribute to grape berry growth and ripening. Valuable contributions to the study of K+ in membrane stabilization, turgor maintenance and phloem transport have allowed us to propose a mechanistic model for the role of this cation in grape berry development.
Keywords: Vitis vinifera; berry; fruit; grapevine; phloem; potassium; ripening; xylem.
Figures
References
-
- Agasse A., Vignault C., Kappel C., Conde C., Gerós H., Delrot S. (2009). “Sugar transport and sugar sensing in grape,” in Grapevine Molecular Physiology and Biotechnology ed. Roubelakis-Angelakis K. A. (Amsterdam: Springer; ) 105–139. 10.1007/978-90-481-2305-6-5 - DOI
-
- Ageorges A., Issaly N., Picaud S., Delrot S., Romieu C. (2000). Identification and functional expression in yeast of a grape berry sucrose carrier. Plant Physiol. Biochem. 38 177–185. 10.1016/S0981-9428(00)00730-0 - DOI
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources