

Review
doi: 10.1111/mpp.12580.
Epub 2017 Oct 11.
Not in your usual Top 10: protists that infect plants and algae
Affiliations
- PMID: 29024322
- PMCID: PMC5772912
- DOI: 10.1111/mpp.12580
Item in Clipboard
Review
Not in your usual Top 10: protists that infect plants and algae
Mol Plant Pathol.
2018 Apr.
Abstract
Fungi, nematodes and oomycetes belong to the most prominent eukaryotic plant pathogenic organisms. Unicellular organisms from other eukaryotic lineages, commonly addressed as protists, also infect plants. This review provides an introduction to plant pathogenic protists, including algae infecting oomycetes, and their current state of research.
Keywords: algae; phytomonas; phytomyxae; plant pathogens; plasmodiophorids; protist; stramenopiles.
© 2017 THE AUTHORS. MOLECULAR PLANT PATHOLOGY PUBLISHED BY BRITISH SOCIETY FOR PLANT PATHOLOGY AND JOHN WILEY & SONS LTD.
Figures
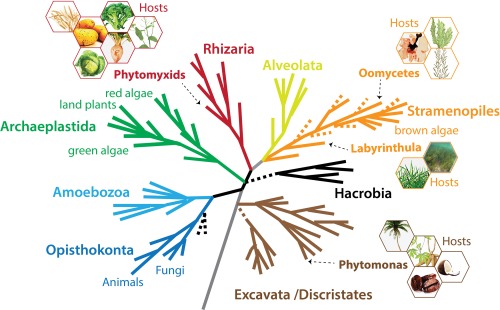
A schematic current eukaryotic tree of life indicating the phylogenetic positions of the eukaryotic plant pathogens outlined in this review. The hexagons show examples of the host species for each pathogen group. The phylogenetic tree was created by S. Baldauf (Uppsala University, Uppsala, Sweden) and reproduced with permission.

Phytomonas sp. and palm infections. (A, B) Scanning electron micrographs of Phytomonas serpens cells in culture (scale bars, 10 and 1 μm). (Courtesy of Martina Tesařová.) (C) Transmission electron micrographs of Phytomonas sp. flagellates in the phloem of coconut palms affected by hartrot. C, companion cell; F, fibre; M, immature metaxylem; P, phloem parenchyma cell; S, sieve elements free of flagellates. (a) Transverse section of a differentiating vascular bundle, showing recently matured sieve elements filled with flagellates (scale bar, 10 μm). (b) Transverse section of the phloem in palm with advanced symptoms (scale bar, 5 μm). (c) Transverse section of a dividing flagellate (scale bar, 0.5 μm). (d) Longitudinal section of a sieve element filled with flagellates. Arrows indicate the kinetoplast DNA (scale bar, 1 μm). (Reproduced from Parthasarathy et al., 1976.) (D) Coconut palms with symptoms of hartrot. (Photograph: Monica L. Elliott, Professor, Plant Pathology, University of Florida, Institute of Food and Agricultural Sciences (UF/IFAS), Gainesville, FL, USA.)
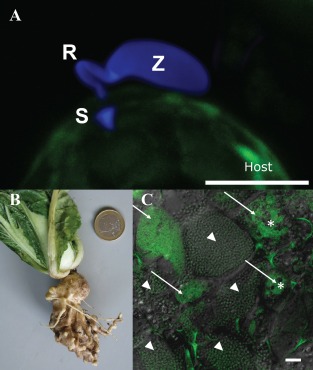
Phytomyxid infection and clubroot. (A) Phytomyxean parasites infect their host via a specialized extrusosome, called a ‘Rohr (R) and Stachel (S)’. The image shows a zoospore (Z) of the phagomyxid Maullinia ectocarpii infecting a female gametophyte of Macrocystis pyrifera (host). The M. ectocarpii spore was stained with calcofluor white and the host is visible via autofluorescence. Bar, 5 µm. (B) Clubroot symptoms on Chinese cabbage. (C) Laser scanning micrograph of Plasmodiophora brassicae resting spores (arrowheads) and plasmodia (arrows) in clubroot tissue. Plasmodia of different ages can be distinguished by the presence of typical vacuoles (asterisks), which disappear when the plasmodia start to differentiate into resting spores. Overlay of a light microscopic image and the signal of a Plasmodiophora‐specific fluorescence in situ hybridization (FISH) probe (green: excitation, 488 nm; emission, 510–550 nm). Bar, 20 µm.

Potato infection by Spongospora subterranea. The potato pathogen Spongospora subterranea infects host tubers, roots and stolons, resulting in the development of powdery scab lesions (A) and galls (B). These usually appear in potato crops 2–3 months after planting, and mature to release sporosori (conglomerations of resting spores). A sporosorus contains 500–1000 resting spores, each containing a primary zoospore (D; bar, 10 µm). Secondary zoospores formed in zoosporangia (C; bar, 20 µm) emerge through root cell walls, disrupting host nutrient and water uptake.
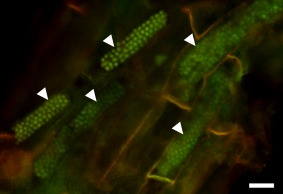
Resting spores of Polymyxa graminis in Poa sp. Resting spores are arranged in typical, long and cylindrical cytosori (arrowheads). The sample was stained with acridine orange, showing the nuclei of the fully developed resting spores. Epifluorescence micrograph obtained using blue excitation with long‐pass emission (Nikon B‐2A filter) allowing for the detection of DNA. Bar, 20 µm.

Labyrinthula and disease symptoms. (A) Single fusiform cells of the unicellular Labyrinthulomycota Labyrinthula protist. (B) Labyrinthula cells emerging from a seagrass leaf on serum seawater agar. Cells move through colonies of self‐generated ectoplasmodic networks or ‘slimeways’, a net‐like tube within which Labyrinthula are able to move. (C) Symptoms of the seagrass wasting disease 4 days following the artificial infection of seagrass blades.

Oomycete phylogeny, Pyropia farming, Pythium porphyrae and Olpidiopsis symptoms. (A) Schematic phylogenetic tree of Oomycetes based on Beakes et al. (2012) indicating the positions of the discussed pathogens of marine algae. (B) Pyropia seaweed harvest on a commercial farm in South Korea (photograph: H. Kim). (C, D) Pyropia blade with lesions caused by Pythium porphyrae (C) and Olpidiopsis (D) infection. Photographs were originally published in Kim et al. (2014) which includes more detailed descriptions of Pyropia diseases.
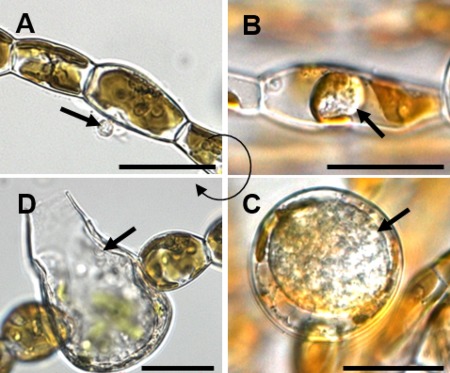
Life cycle of Eurychasma dicksonii in its brown algal host Ectocarpus siliculosus. (A) A spore (arrow) attaches to the algal surface and injects its content into the host. (B) Within the algal cytoplasm, the Eu. dicksonii thallus (arrow) develops which, at the early stage of infection, is unwalled. (C) Later, the pathogen thallus (arrow) has a cell wall and causes hypertrophic expansion of the algal host cell. (D) At the final stage, the complete thallus differentiates into a sporangium from which motile zoospores (arrow) are produced, completing the life cycle of the pathogen. Scale bars equal to 25 μm. (Figure reproduced from Strittmatter et al. 2016.)
References
-
- Adl, S.M. , Simpson, A.G.B. , Lane, C.E. , Lukes, J. , Bass, D. , Bowser, S.S. , Brown, M.W. , Burki, F. , Dunthorn, M. , Hampl, V. , Heiss, A. , Hoppenrath, M. , Lara, E. , Le Gall, L. , Lynn, D.H. , McManus, H. , Mitchell, E.A. , Mozley‐Stanridge, S.E. , Parfrey, L.W. , Pawlowski, J. , Rueckert, S. , Shadwick, L. , Schoch, C.L. , Smirnov, A. and Spiegel, F.W. (2012) The revised classification of eukaryotes. J. Eukaryot. Microbiol. 59, 429–493. - PMC - PubMed
-
- Agarwal, A. , Kaul, V. , Faggian, R. , Rookes, J.E. , Ludwig‐Muller, J. and Cahill, D.M. (2011) Analysis of global host gene expression during the primary phase of the Arabidopsis thaliana–Plasmodiophora brassicae interaction. Funct. Plant Biol. 38, 462–478. - PubMed
-
- Aist, J.R. and Williams, P.H. (1971) The cytology and kinetics of cabbage root hair penetration by Plasmodiophora brassicae . Can. J. Bot. 49, 2023–2034.
-
- Amano, H. , Suginaga, R. , Arashima, K. and Noda, H. (1995) Immunological detection of the fungal parasite, Pythium sp. – the causative organism of red rot disease in Porphyra‐yezoensis . J. Appl. Phycol. 7, 53–58.
-
- Amon, J.P. (1978) Thraustochytrids and labyrinthulids of terrestrial, aquatic and hypersaline environments of the Great Salt Lake, USA. Mycologia, 70, 1299–1301.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
