

The heart of PD: Lewy body diseases as neurocardiologic disorders
- PMID: 29030055
- PMCID: PMC10712237
- DOI: 10.1016/j.brainres.2017.09.033
The heart of PD: Lewy body diseases as neurocardiologic disorders
Abstract
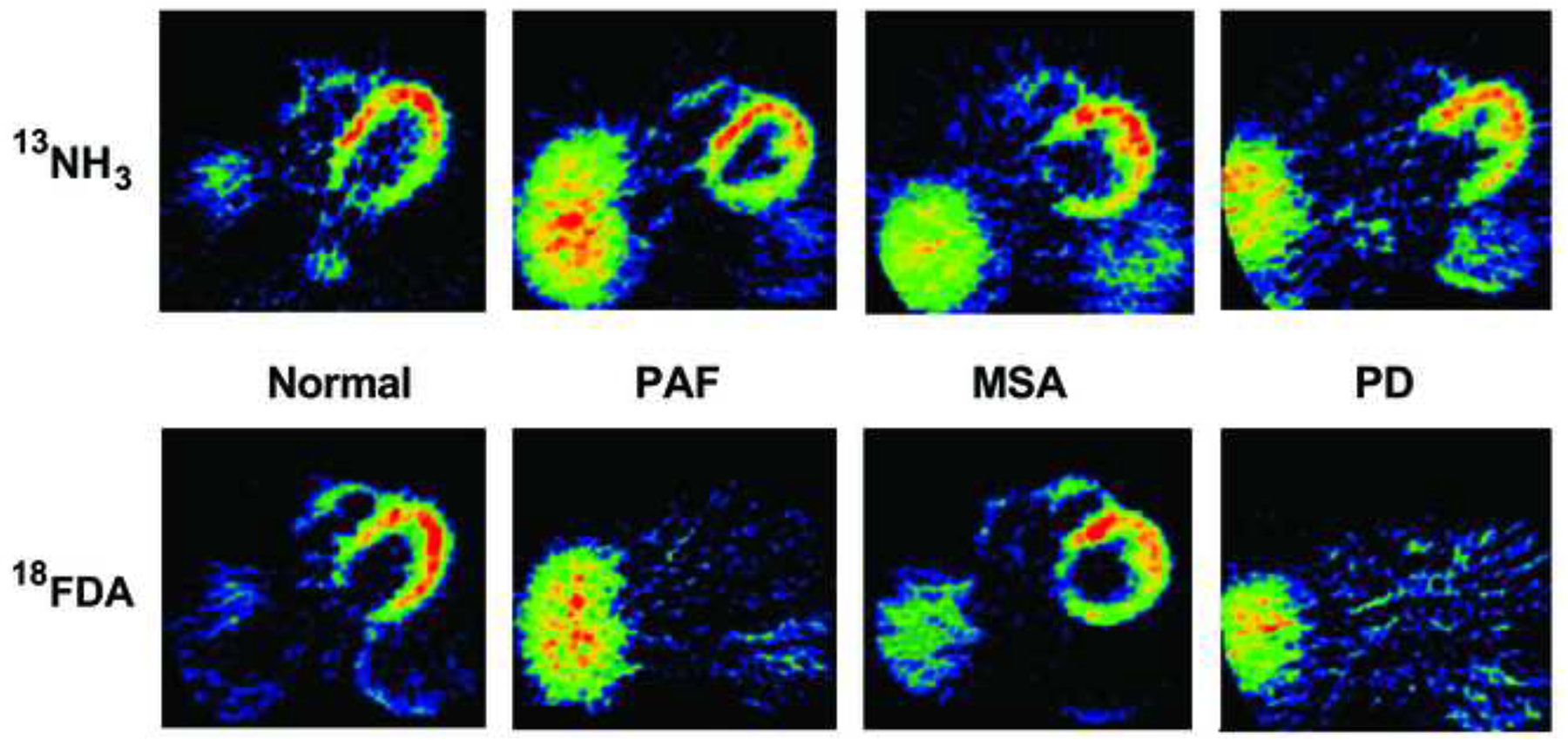
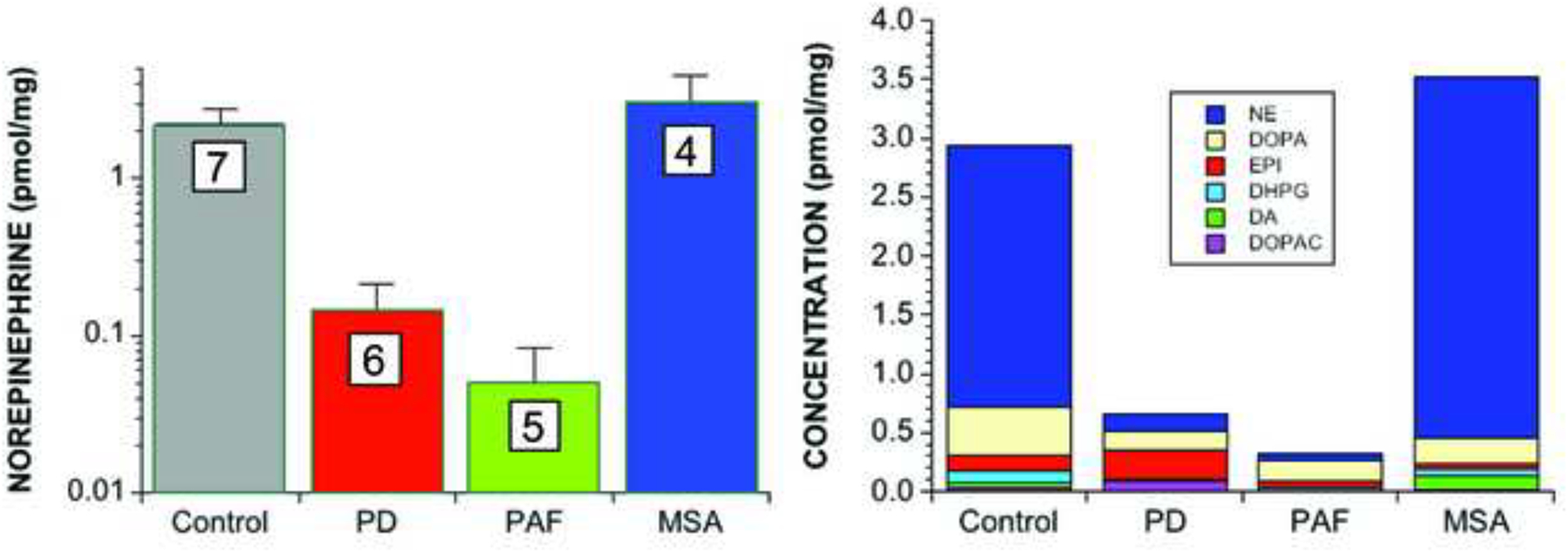
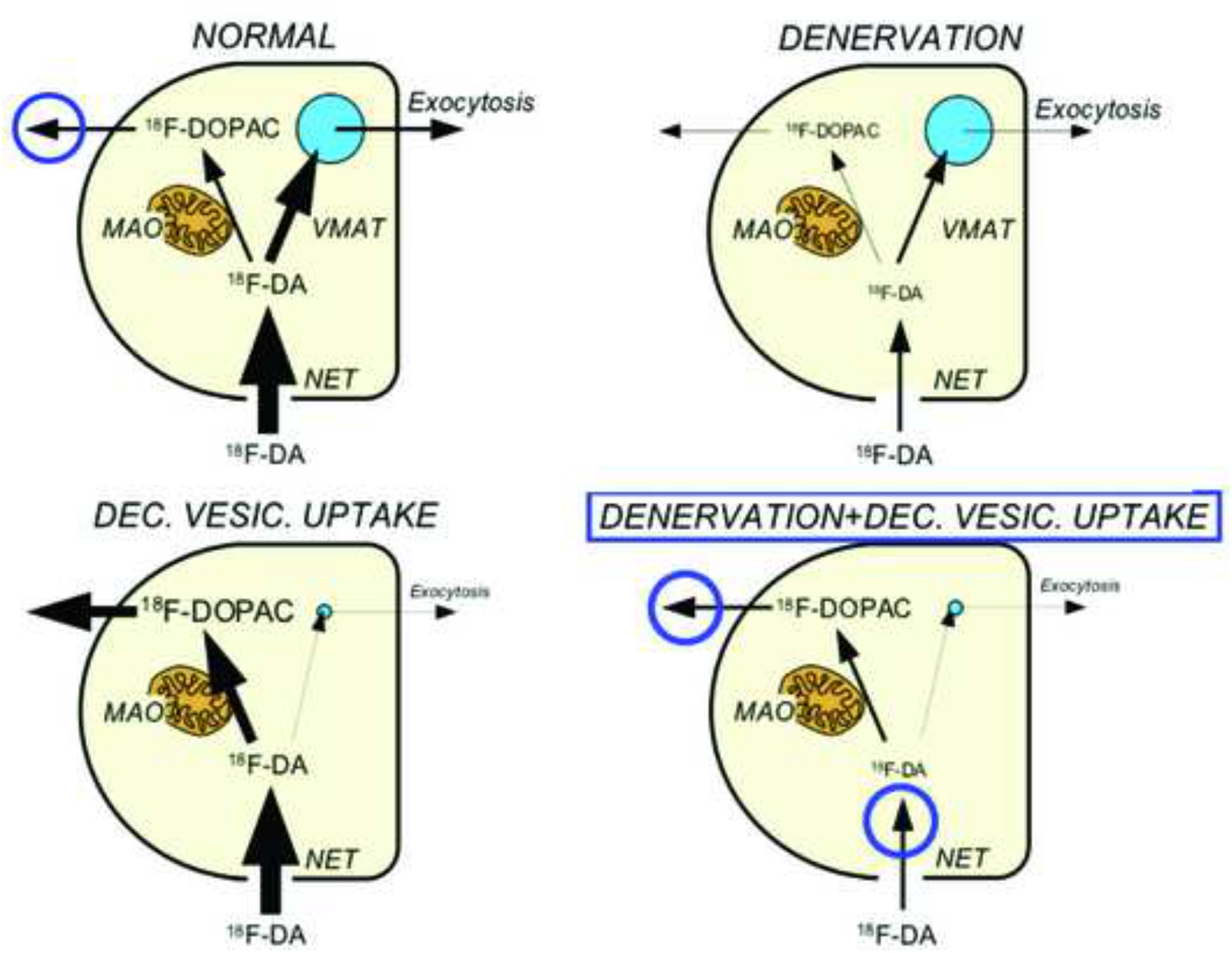
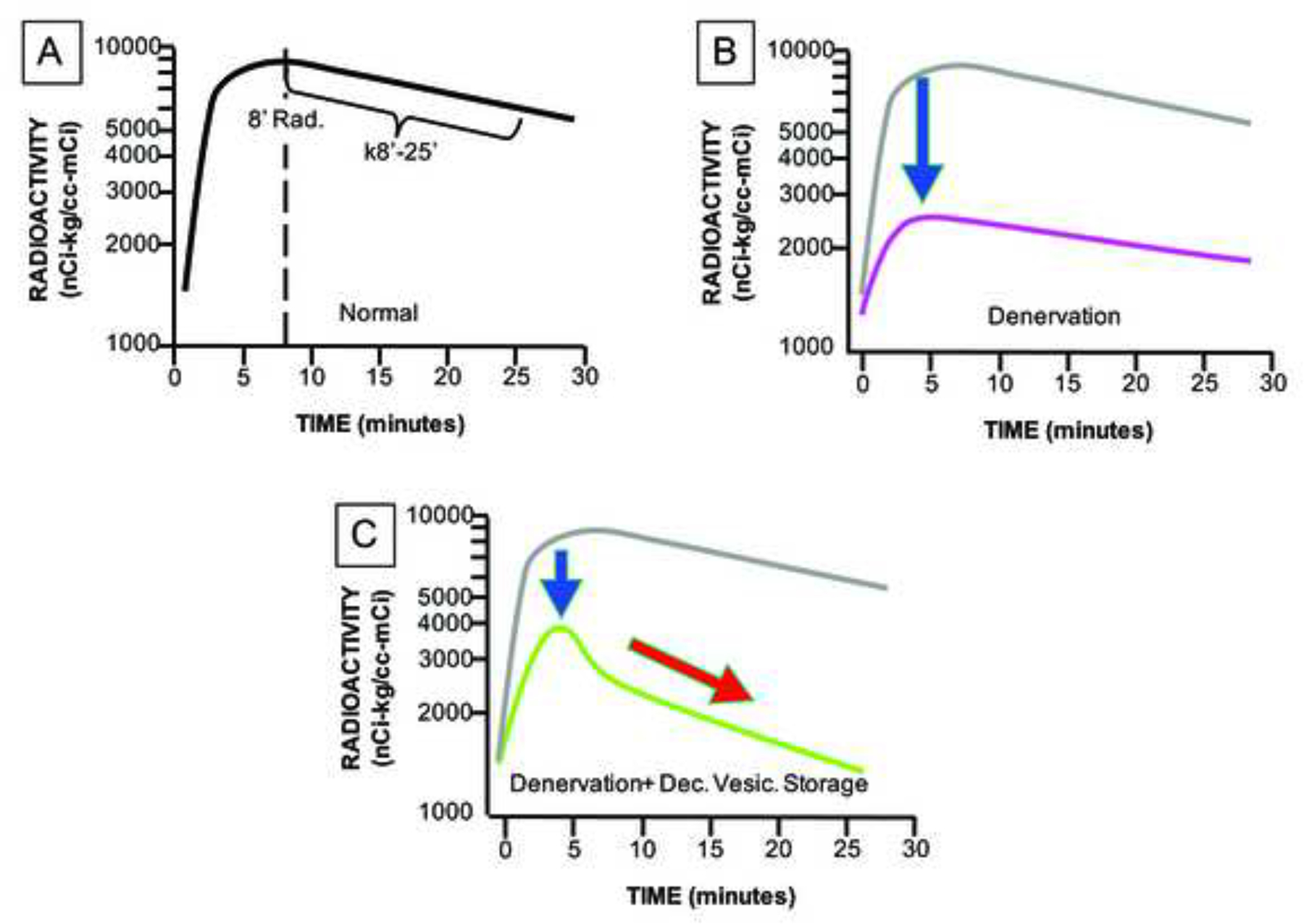
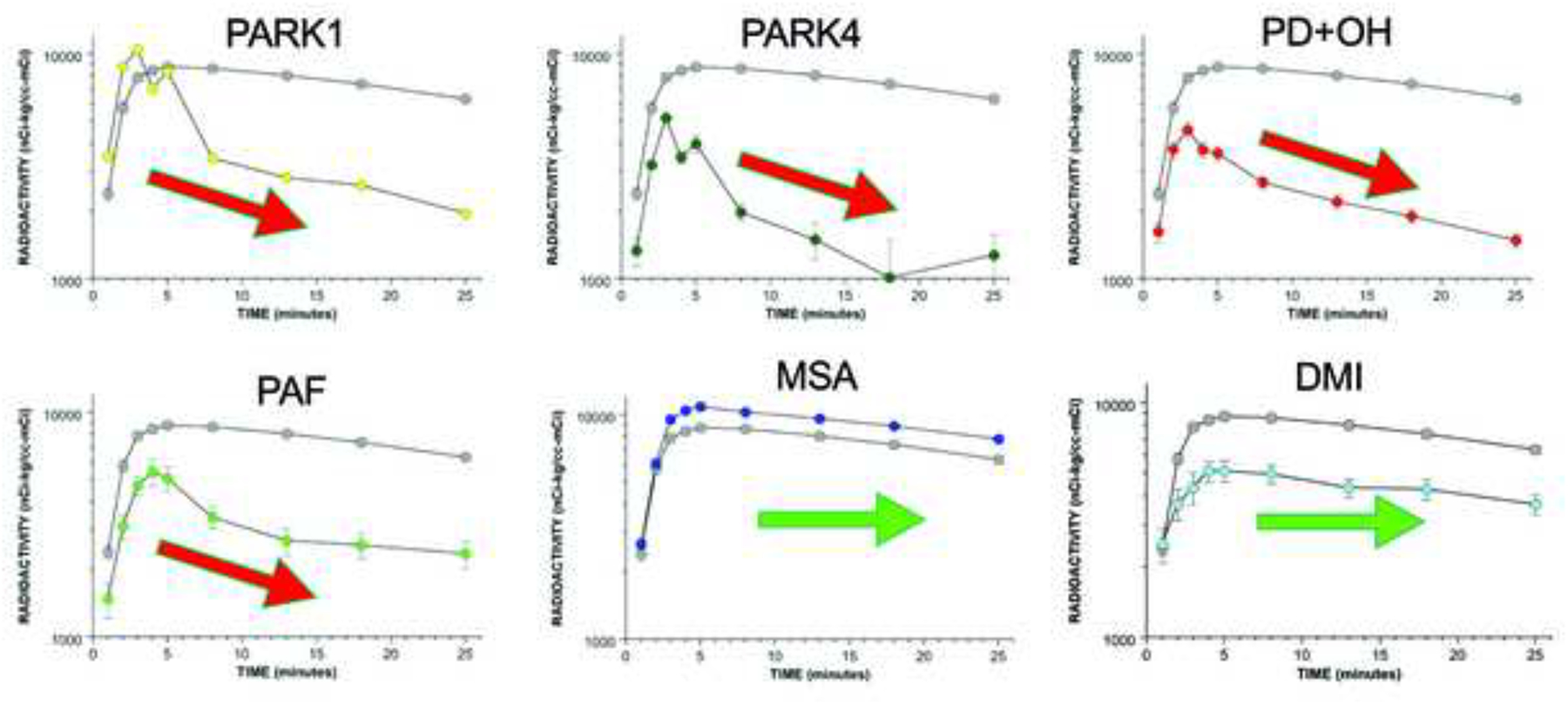
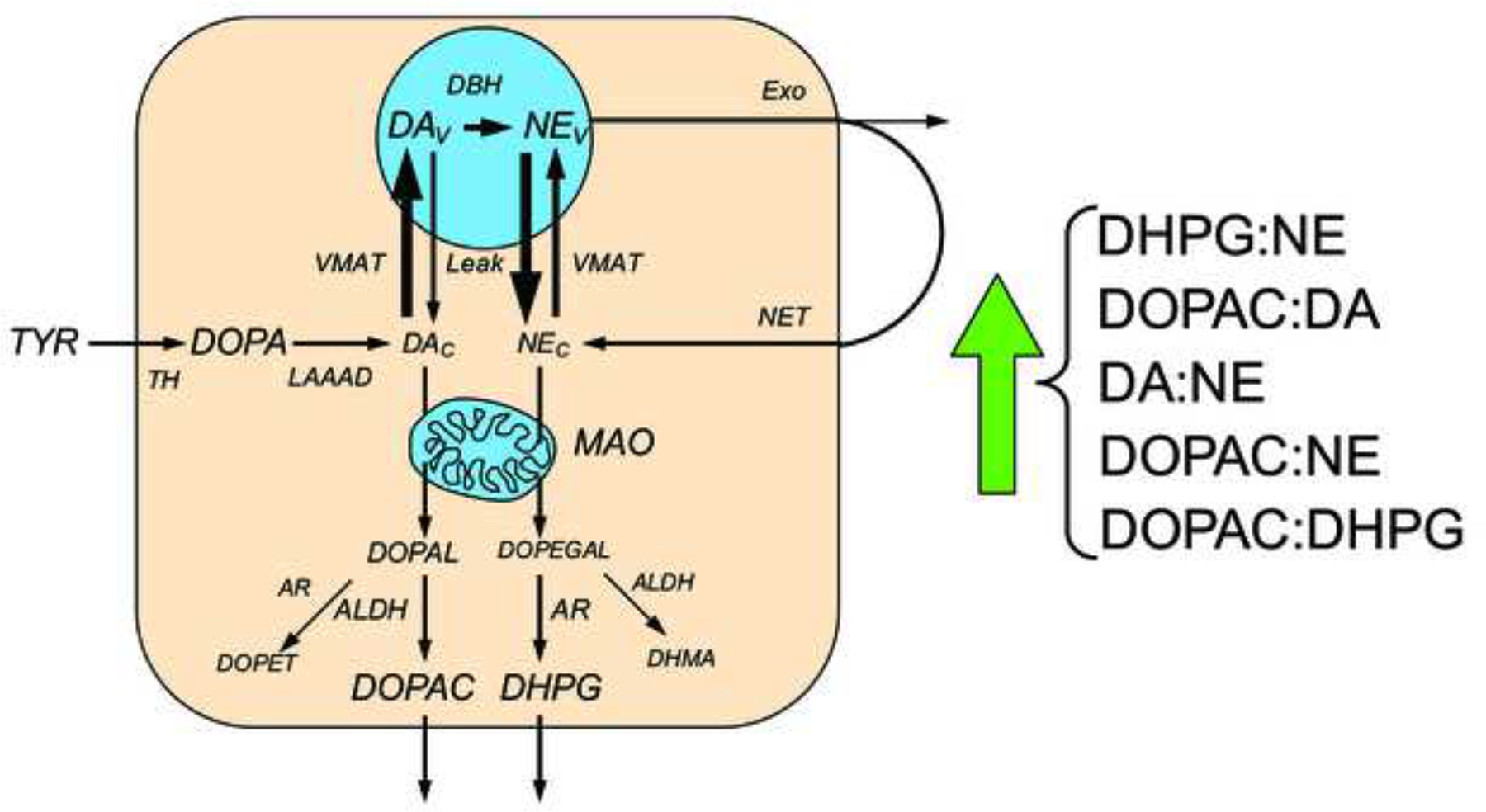
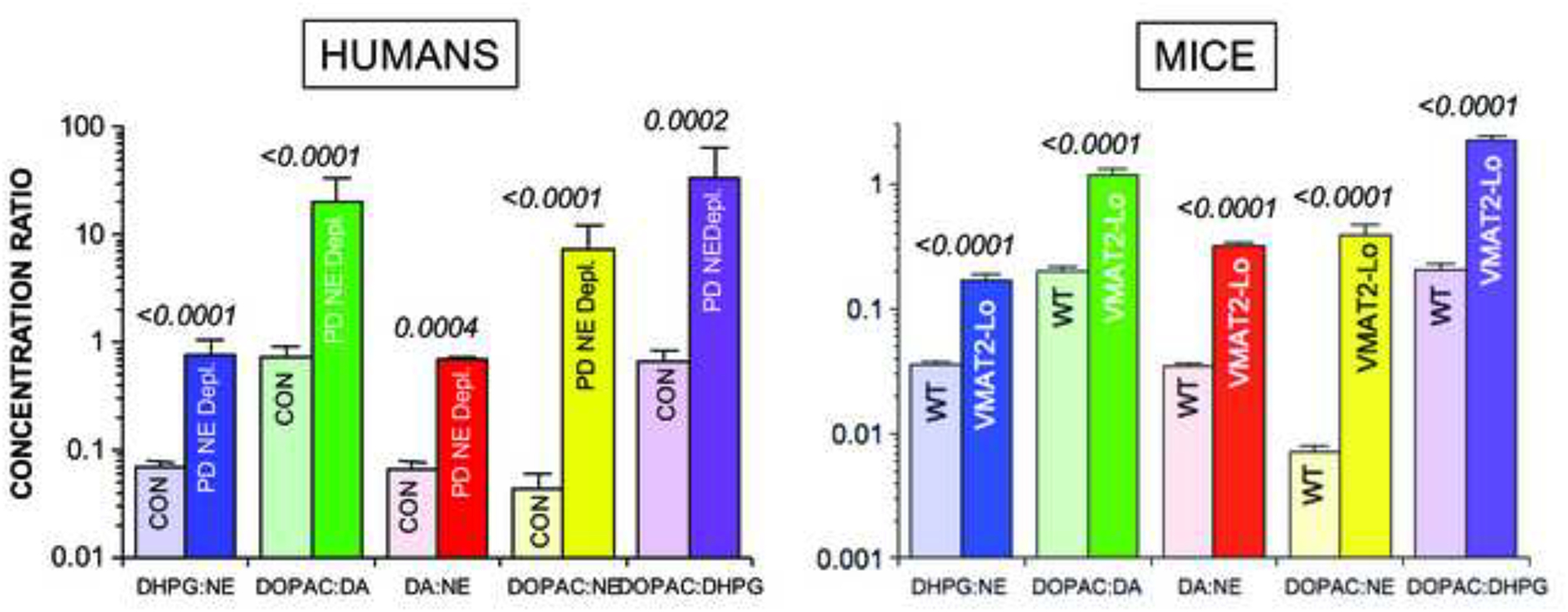
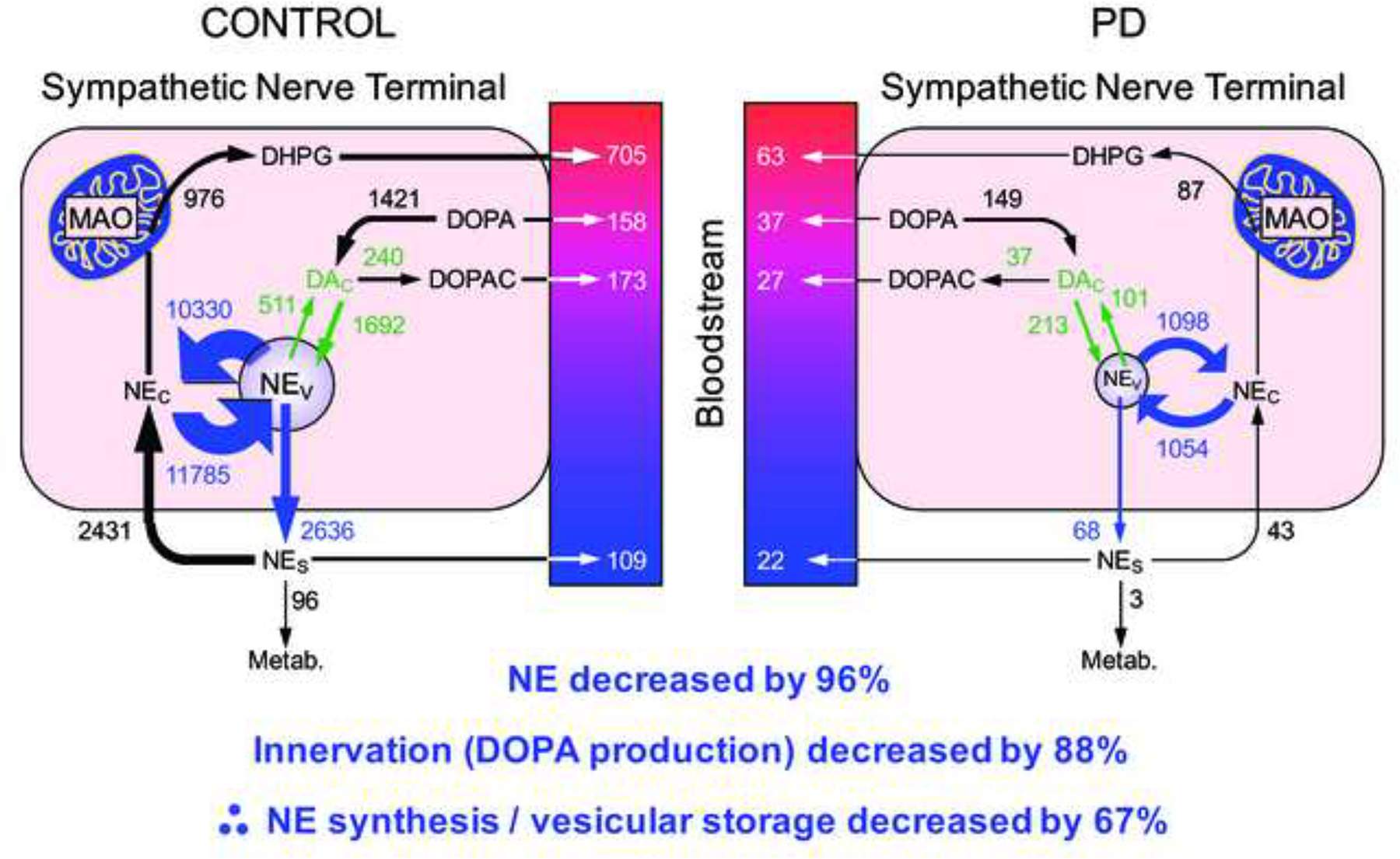
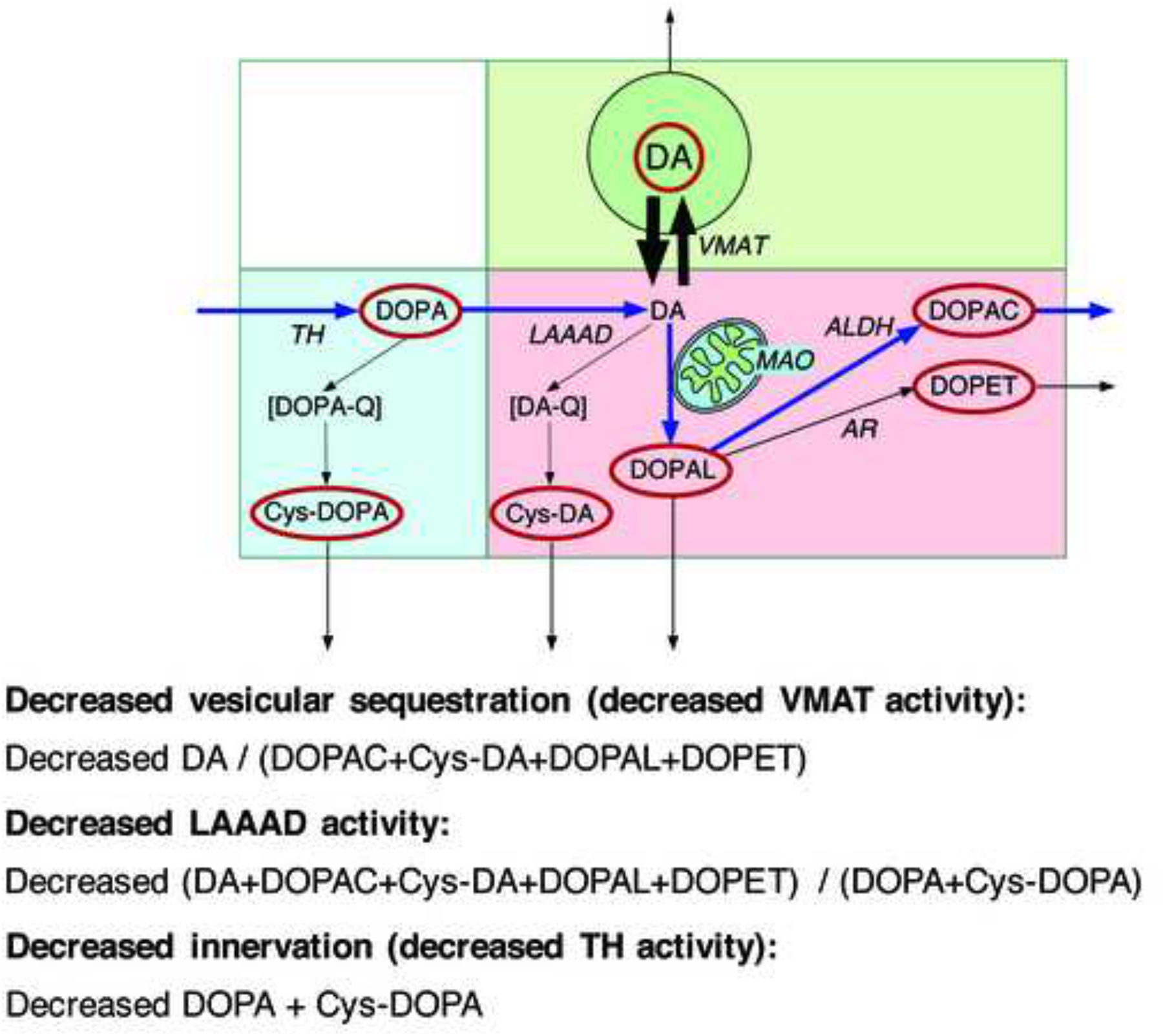
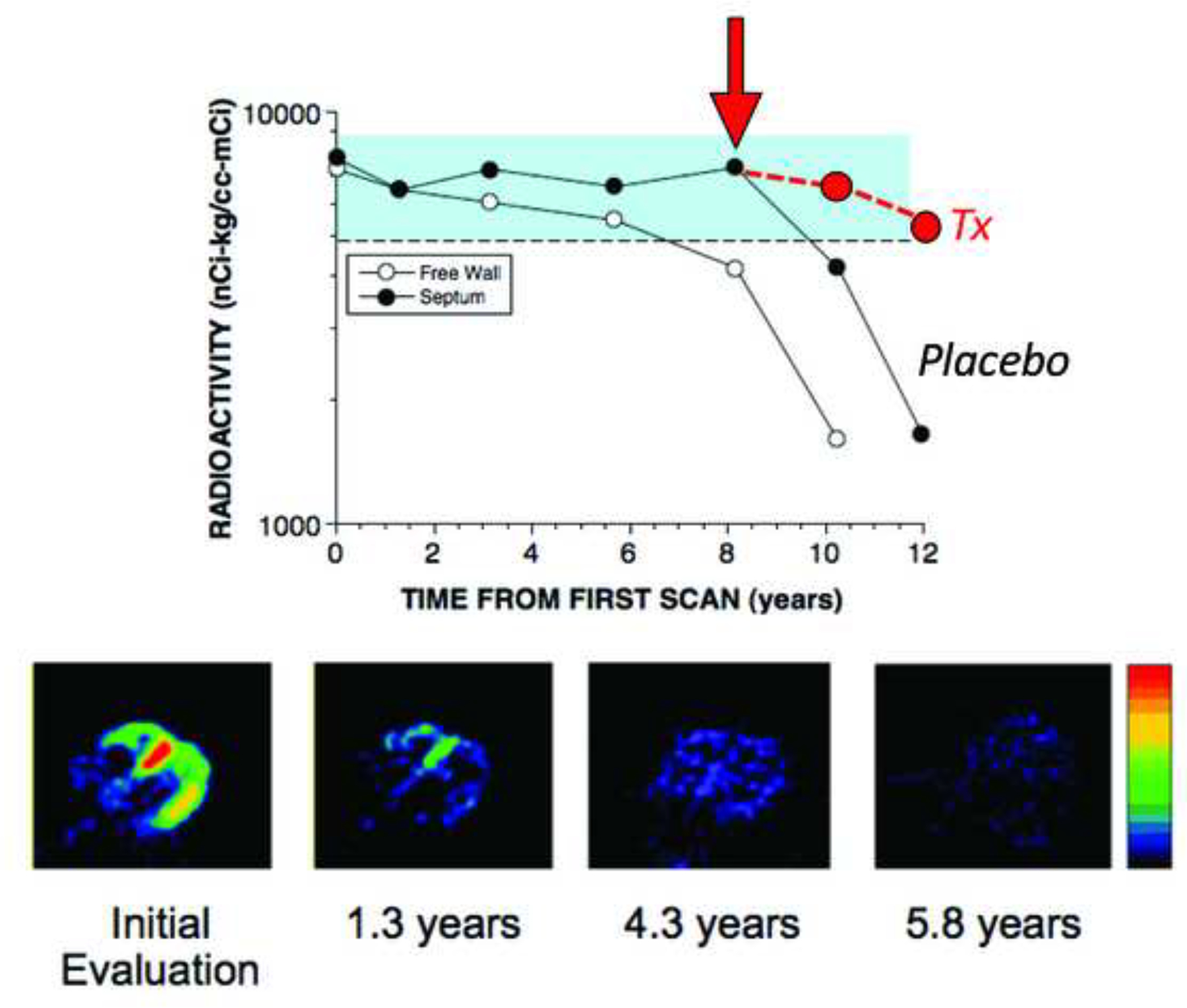
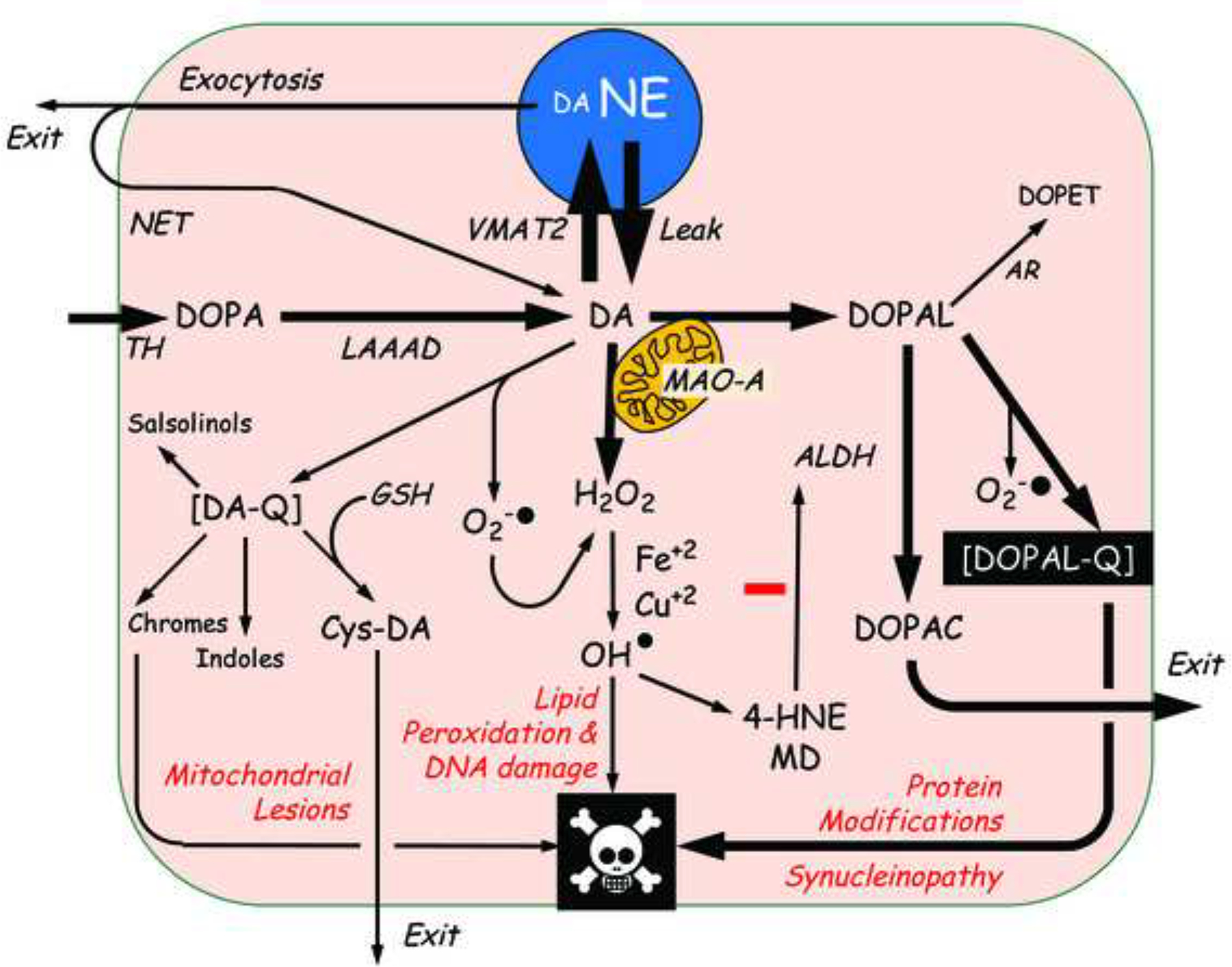
This review provides an update about cardiac sympathetic denervation in Lewy body diseases. The family of Lewy body diseases includes Parkinson's disease (PD), pure autonomic failure (PAF), and dementia with Lewy bodies (DLB). All three feature intra-neuronal cytoplasmic deposits of the protein, alpha-synuclein. Multiple system atrophy (MSA), the parkinsonian form of which can be difficult to distinguish from PD with orthostatic hypotension, involves glial cytoplasmic inclusions that contain alpha-synuclein. By now there is compelling neuroimaging, neuropathologic, and neurochemical evidence for cardiac sympathetic denervation in Lewy body diseases. In addition to denervation, there is decreased storage of catecholamines in the residual terminals. The degeneration develops in a centripetal, retrograde, "dying back" sequence. Across synucleinopathies the putamen and cardiac catecholaminergic lesions seem to occur independently of each other, whereas non-motor aspects of PD (e.g., anosmia, dementia, REM behavior disorder, OH) are associated with each other and with cardiac sympathetic denervation. Cardiac sympathetic denervation can be caused by synucleinopathy in inherited PD. According to the catecholaldehyde hypothesis, 3,4-dihydroxyphenylacetaldehyde (DOPAL), an intermediary metabolite of dopamine, causes or contributes to the death of catecholamine neurons, especially by interacting with proteins such as alpha-synuclein. DOPAL oxidizes spontaneously to DOPAL-quinone, which probably converts alpha-synuclein to its toxic oligomeric form. Decreasing DOPAL production and oxidation might slow the neurodegenerative process. Tracking cardiac sympathetic innervation over time could be the basis for a proof of principle experimental therapeutics trial targeting DOPAL.
Keywords: DOPAL; Dopamine; Norepinephrine; Parkinson’s disease; Sympathetic nervous system.
Published by Elsevier B.V.
Conflict of interest statement
CONFLICT OF INTEREST
The authors have no conflicts of interest to disclose.
Figures
References
-
- Braak H, Ghebremedhin E, Rub U, Bratzke H, Del Tredici K, 2004. Stages in the development of Parkinson’s disease-related pathology. Cell Tissue Res. 318, 121–134. - PubMed
-
- Burke WJ, Li SW, Williams EA, Nonneman R, Zahm DS, 2003. 3,4-Dihydroxyphenylacetaldehyde is the toxic dopamine metabolite in vivo: implications for Parkinson’s disease pathogenesis. Brain Res. 989, 205–13. - PubMed
-
- Burke WJ, Kumar VB, Pandey N, Panneton WM, Gan Q, Franko MW, O’Dell M, Li SW, Pan Y, Chung HD, Galvin JE, 2008. Aggregation of alpha-synuclein by DOPAL, the monoamine oxidase metabolite of dopamine. Acta Neuropathol. 115, 193–203. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
