

Top-Down Control of Visual Attention by the Prefrontal Cortex. Functional Specialization and Long-Range Interactions
- PMID: 29033784
- PMCID: PMC5626849
- DOI: 10.3389/fnins.2017.00545
Top-Down Control of Visual Attention by the Prefrontal Cortex. Functional Specialization and Long-Range Interactions
Abstract
The ability to select information that is relevant to current behavioral goals is the hallmark of voluntary attention and an essential part of our cognition. Attention tasks are a prime example to study at the neuronal level, how task related information can be selectively processed in the brain while irrelevant information is filtered out. Whereas, numerous studies have focused on elucidating the mechanisms of visual attention at the single neuron and population level in the visual cortices, considerably less work has been devoted to deciphering the distinct contribution of higher-order brain areas, which are known to be critical for the employment of attention. Among these areas, the prefrontal cortex (PFC) has long been considered a source of top-down signals that bias selection in early visual areas in favor of the attended features. Here, we review recent experimental data that support the role of PFC in attention. We examine the existing evidence for functional specialization within PFC and we discuss how long-range interactions between PFC subregions and posterior visual areas may be implemented in the brain and contribute to the attentional modulation of different measures of neural activity in visual cortices.
Keywords: executive control; feature attention; long-range interactions; oscillatory synchrony; prefrontal cortex; review; spatial attention; visual cortex.
Figures
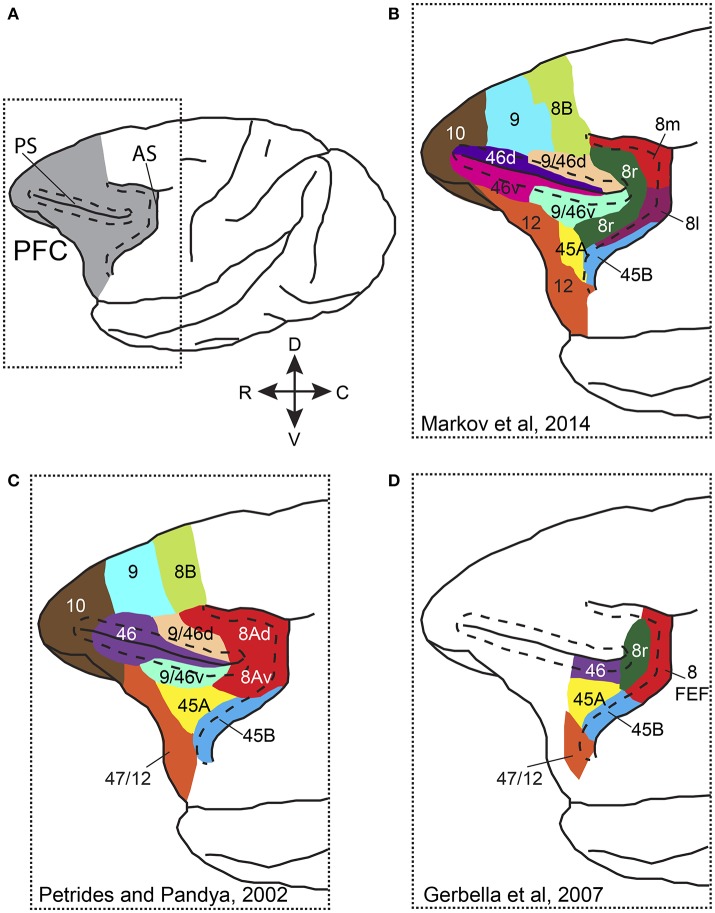
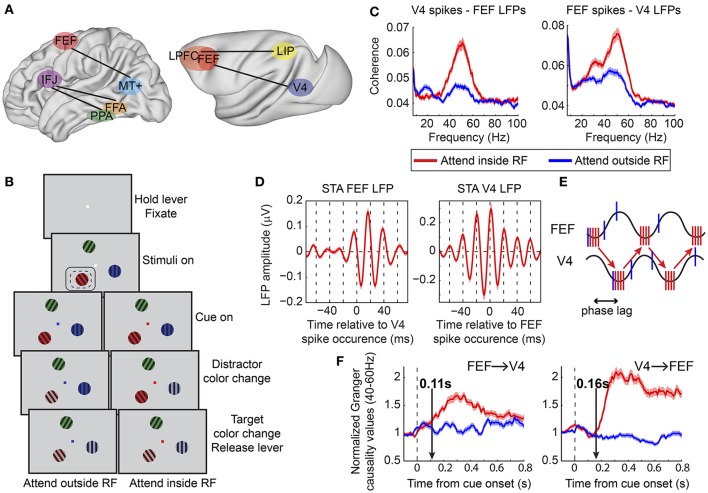
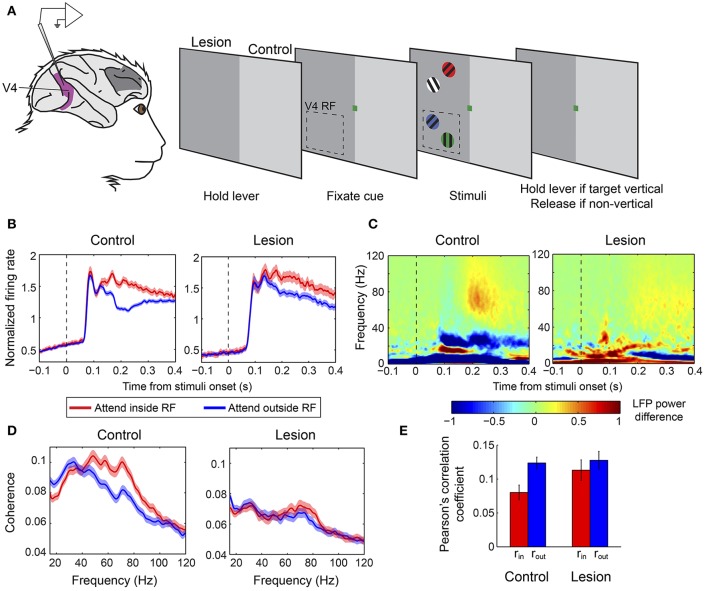
References
-
- Ardid S., Wang X. J., Gomez-Cabrero D., Compte A. (2010). Reconciling coherent oscillation with modulation of irregular spiking activity in selective attention: gamma-range synchronization between sensory and executive cortical areas. J. Neurosci. 30, 2856–2870. 10.1523/JNEUROSCI.4222-09.2010 - DOI - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous