

Spontaneous and Evoked Activity from Murine Ventral Horn Cultures on Microelectrode Arrays
- PMID: 29033792
- PMCID: PMC5626830
- DOI: 10.3389/fncel.2017.00304
Spontaneous and Evoked Activity from Murine Ventral Horn Cultures on Microelectrode Arrays
Abstract
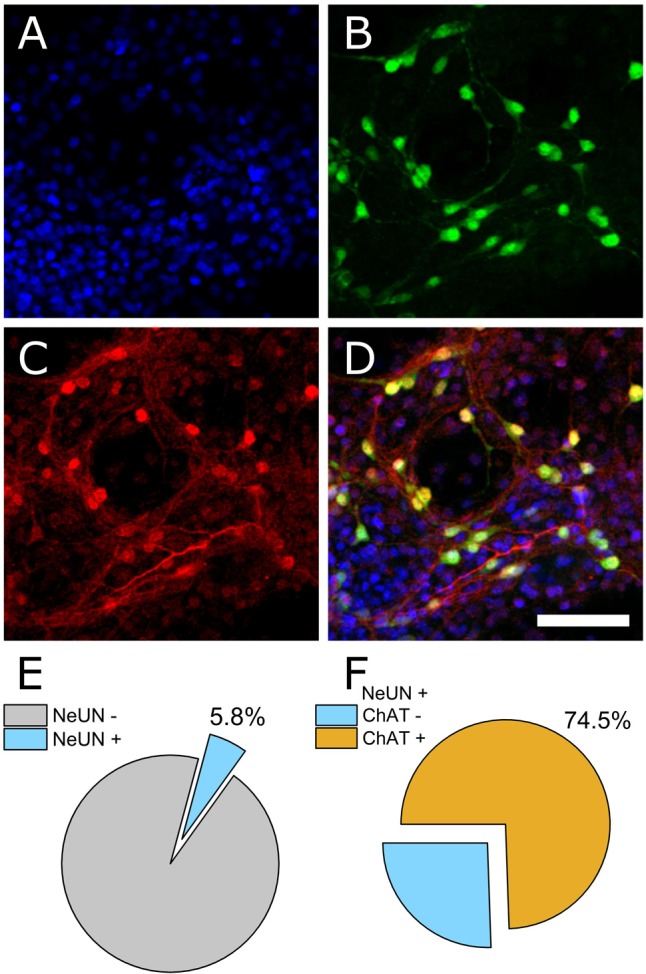
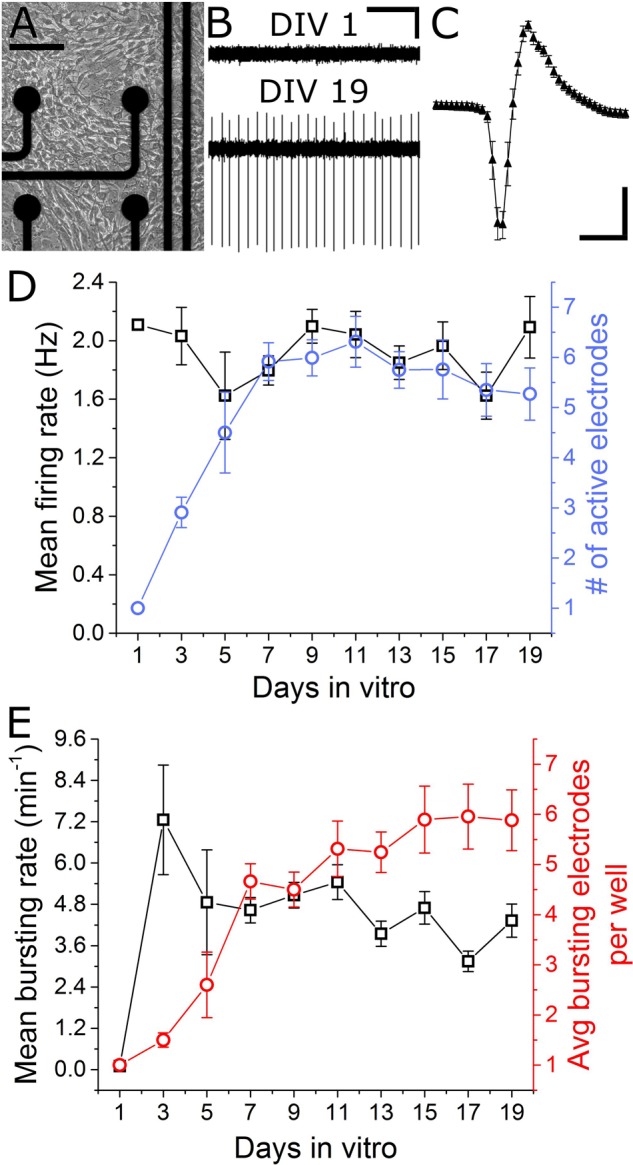
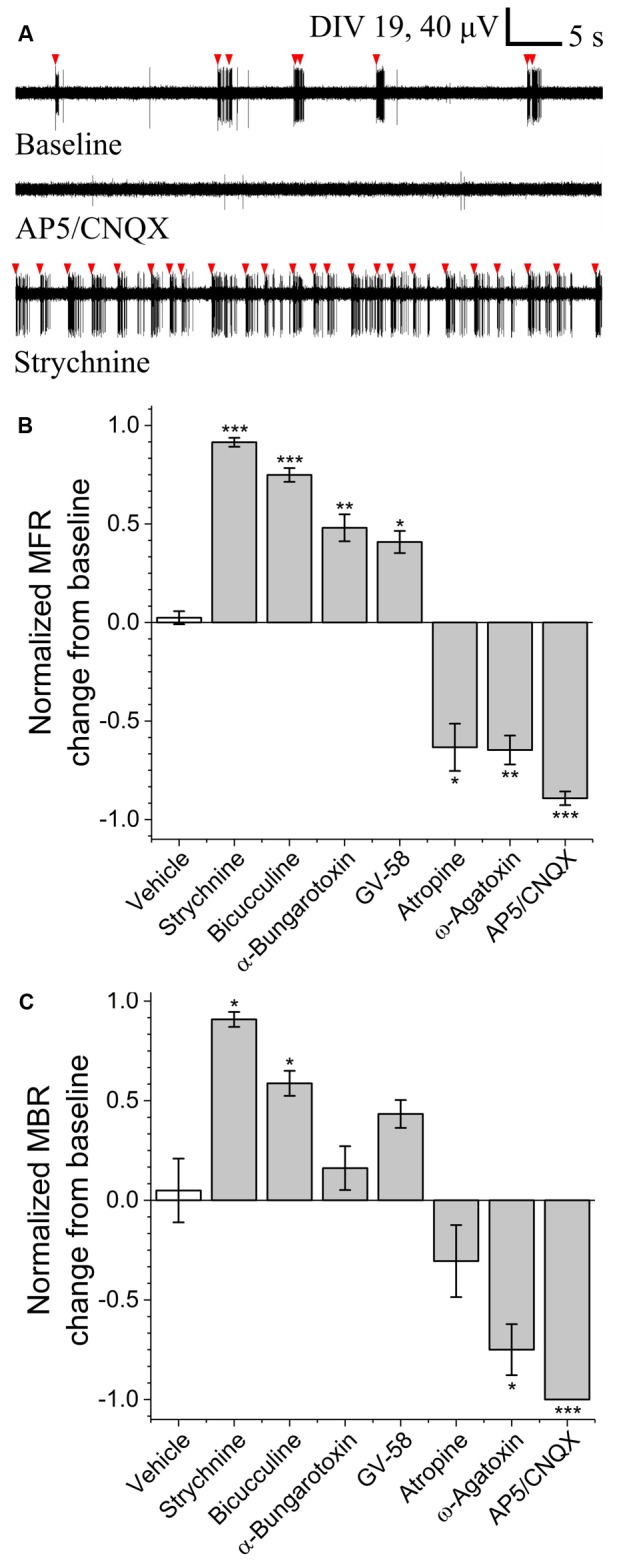
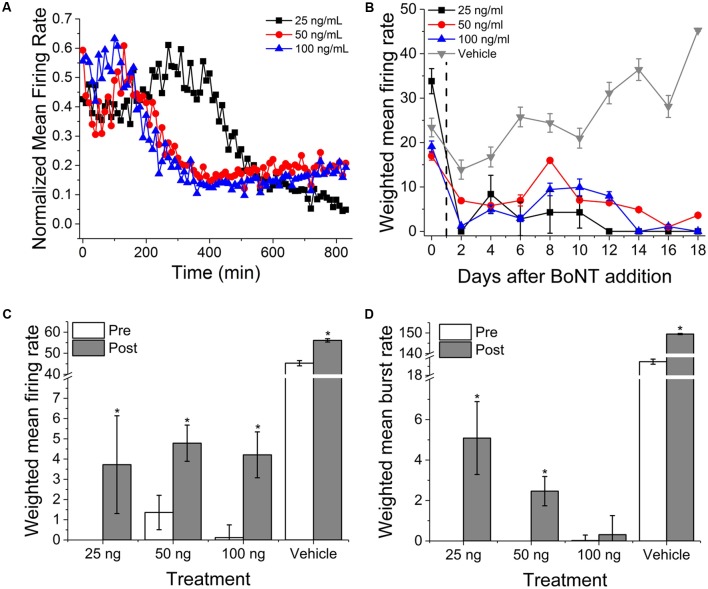
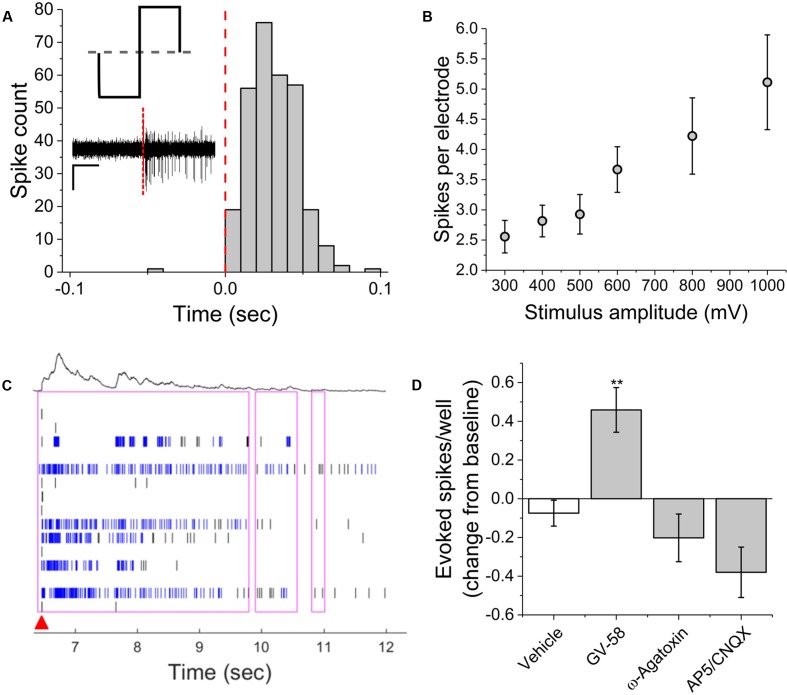
Motor neurons are the site of action for several neurological disorders and paralytic toxins, with cell bodies located in the ventral horn (VH) of the spinal cord along with interneurons and support cells. Microelectrode arrays (MEAs) have emerged as a high content assay platform for mechanistic studies and drug discovery. Here, we explored the spontaneous and evoked electrical activity of VH cultures derived from embryonic mouse spinal cord on multi-well plates of MEAs. Primary VH cultures from embryonic day 15-16 mice were characterized by expression of choline acetyltransferase (ChAT) by immunocytochemistry. Well resolved, all-or-nothing spontaneous spikes with profiles consistent with extracellular action potentials were observed after 3 days in vitro, persisting with consistent firing rates until at least day in vitro 19. The majority of the spontaneous activity consisted of tonic firing interspersed with coordinated bursting across the network. After 5 days in vitro, spike activity was readily evoked by voltage pulses where a minimum amplitude and duration required for excitation was 300 mV and 100 μs/phase, respectively. We characterized the sensitivity of spontaneous and evoked activity to a host of pharmacological agents including AP5, CNQX, strychnine, ω-agatoxin IVA, and botulinum neurotoxin serotype A (BoNT/A). These experiments revealed sensitivity of the cultured VH to both agonist and antagonist compounds in a manner consistent with mature tissue derived from slices. In the case of BoNT/A, we also demonstrated intoxication persistence over an 18-day period, followed by partial intoxication recovery induced by N- and P/Q-type calcium channel agonist GV-58. In total, our findings suggest that VH cultures on multi-well MEA plates may represent a moderate throughput, high content assay for performing mechanistic studies and for screening potential therapeutics pertaining to paralytic toxins and neurological disorders.
Keywords: BoNT/A; electrical stimulation; in vitro models; microelectrode array; motor neuron; neural activity modulation; neural activity recording; ventral horn.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources