

mTORC1 stimulates phosphatidylcholine synthesis to promote triglyceride secretion
- PMID: 29035283
- PMCID: PMC5663357
- DOI: 10.1172/JCI96036
mTORC1 stimulates phosphatidylcholine synthesis to promote triglyceride secretion
Abstract
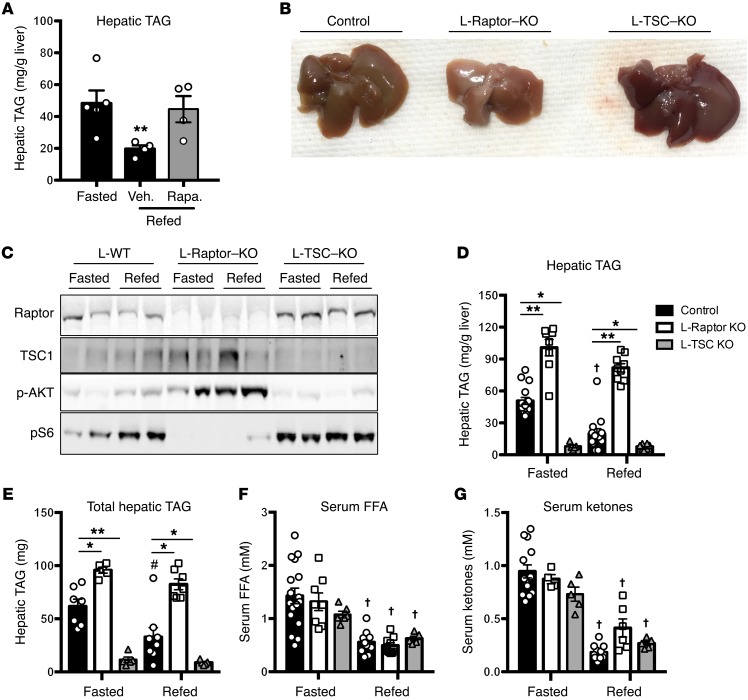
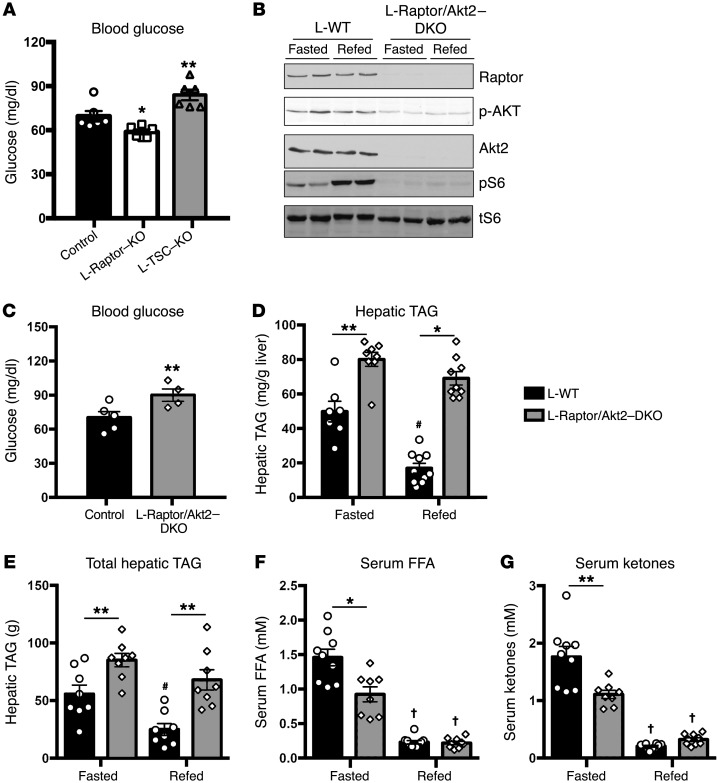
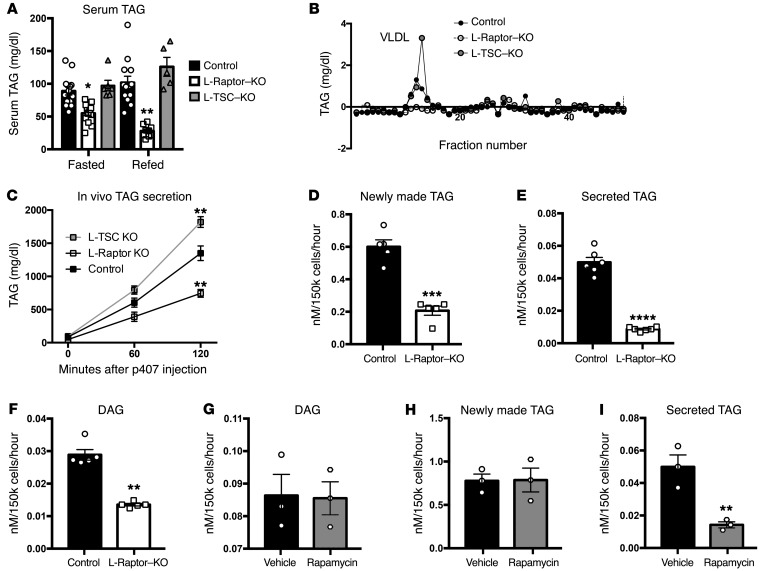
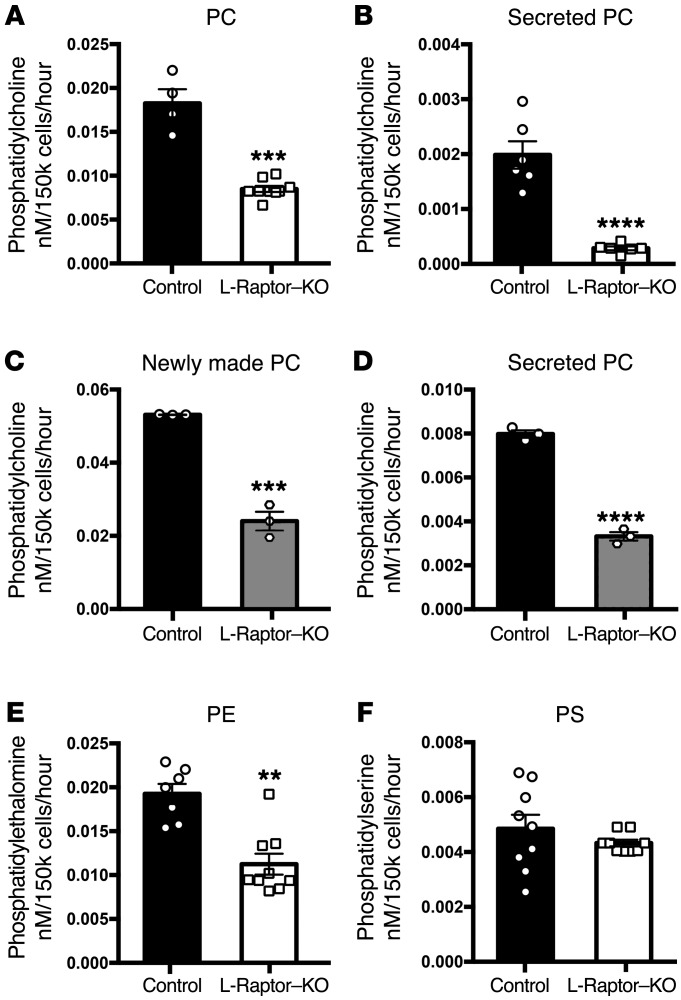
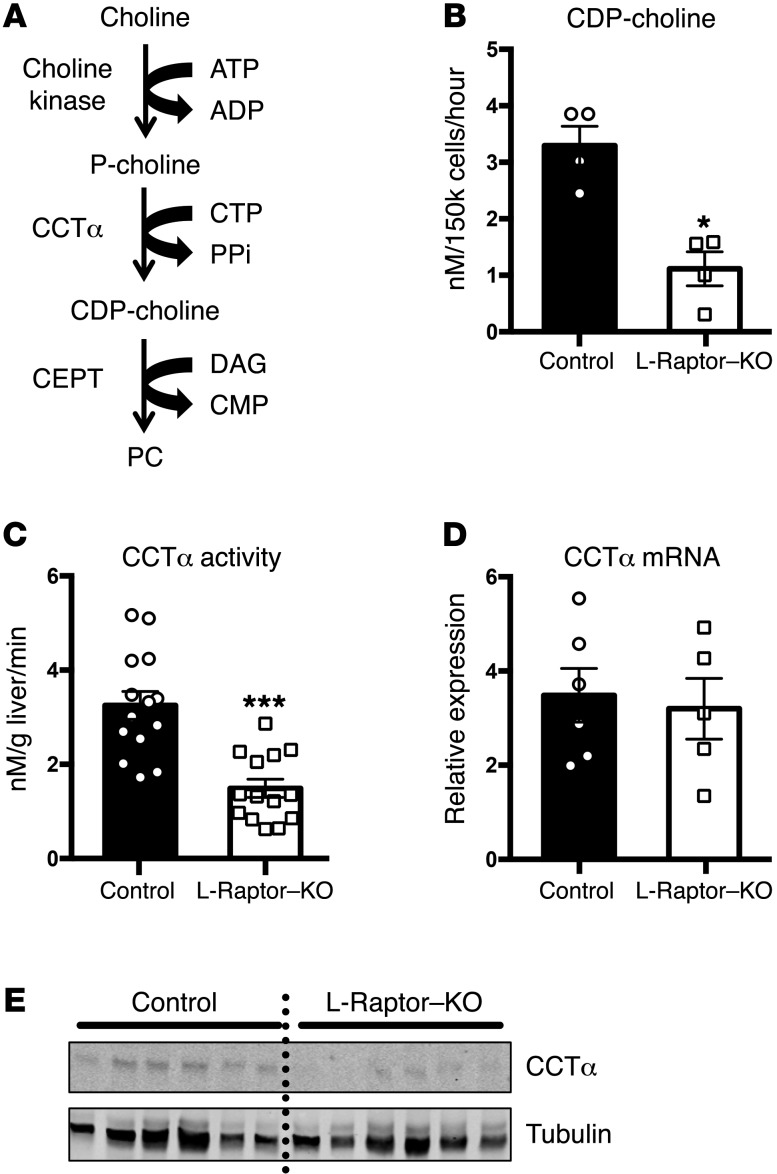
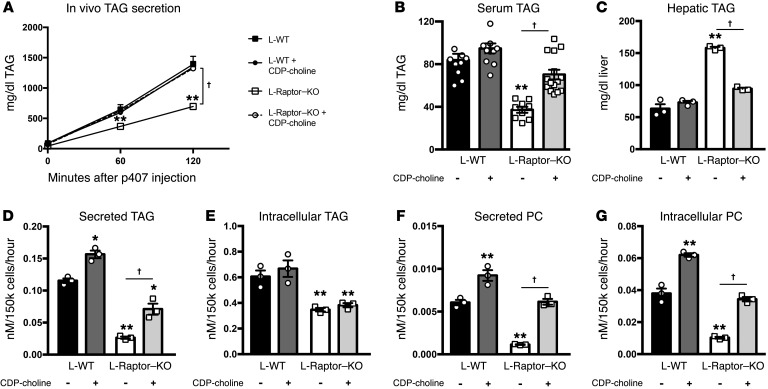
Liver triacylglycerol (TAG) synthesis and secretion are closely linked to nutrient availability. After a meal, hepatic TAG formation from fatty acids is decreased, largely due to a reduction in circulating free fatty acids (FFA). Despite the postprandial decrease in FFA-driven esterification and oxidation, VLDL-TAG secretion is maintained to support peripheral lipid delivery and metabolism. The regulatory mechanisms underlying the postprandial control of VLDL-TAG secretion remain unclear. Here, we demonstrated that the mTOR complex 1 (mTORC1) is essential for this sustained VLDL-TAG secretion and lipid homeostasis. In murine models, the absence of hepatic mTORC1 reduced circulating TAG, despite hepatosteatosis, while activation of mTORC1 depleted liver TAG stores. Additionally, mTORC1 promoted TAG secretion by regulating phosphocholine cytidylyltransferase α (CCTα), the rate-limiting enzyme involved in the synthesis of phosphatidylcholine (PC). Increasing PC synthesis in mice lacking mTORC1 rescued hepatosteatosis and restored TAG secretion. These data identify mTORC1 as a major regulator of phospholipid biosynthesis and subsequent VLDL-TAG secretion, leading to increased postprandial TAG secretion.
Conflict of interest statement
Figures
References
-
- Adamovich Y, Aviram R, Asher G. The emerging roles of lipids in circadian control. Biochim Biophys Acta. 2015;1851(8):1017–1025. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
