

The role of the ER stress-response protein PERK in rhodopsin retinitis pigmentosa
- PMID: 29036441
- PMCID: PMC5868081
- DOI: 10.1093/hmg/ddx370
The role of the ER stress-response protein PERK in rhodopsin retinitis pigmentosa
Abstract
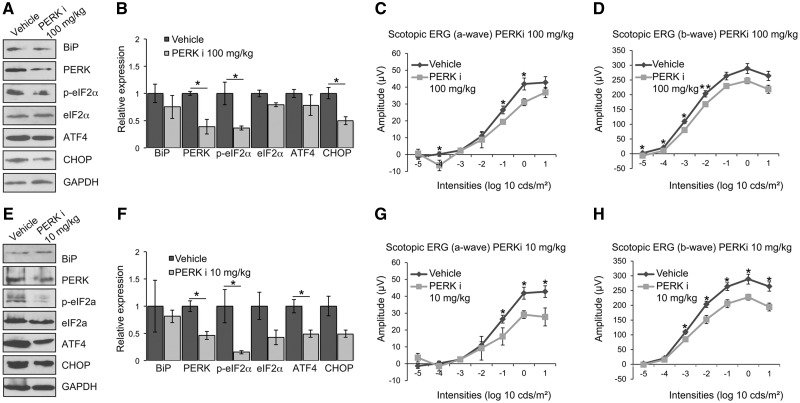
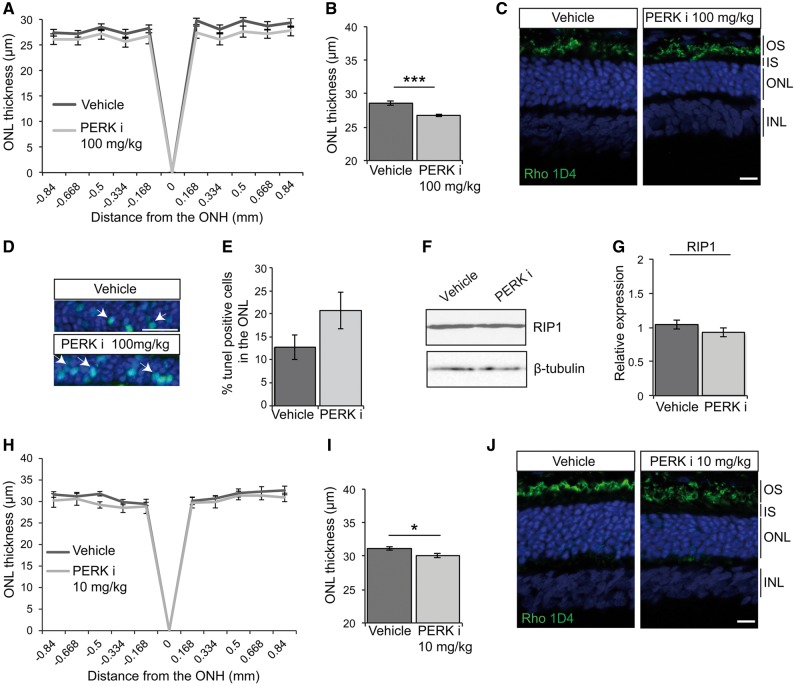
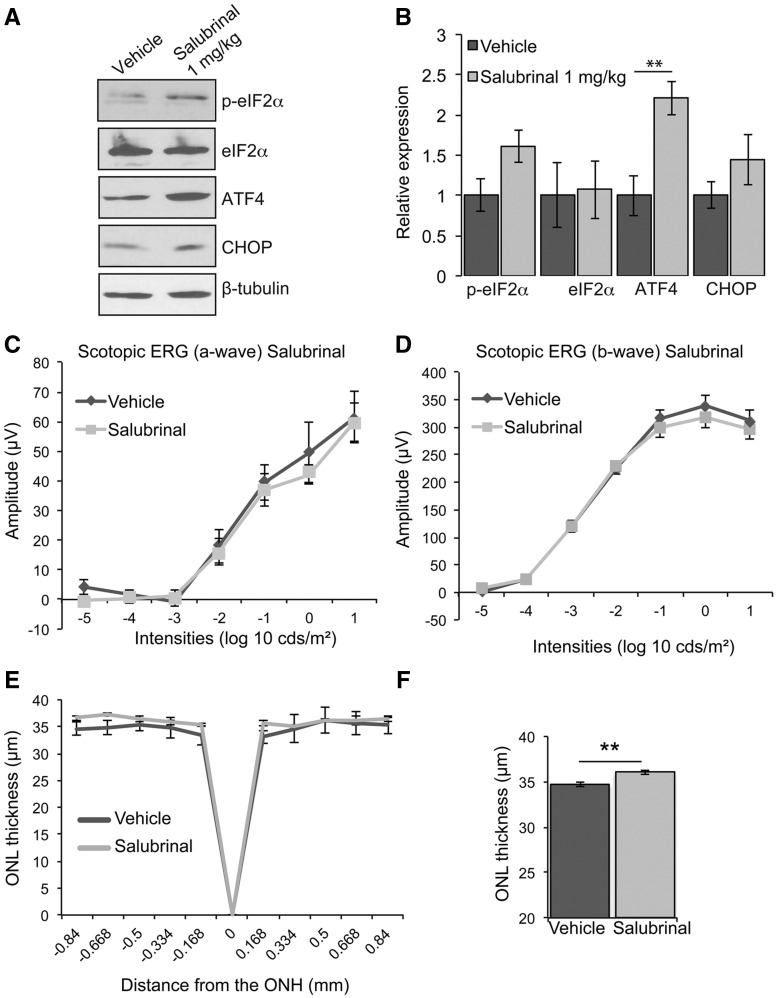
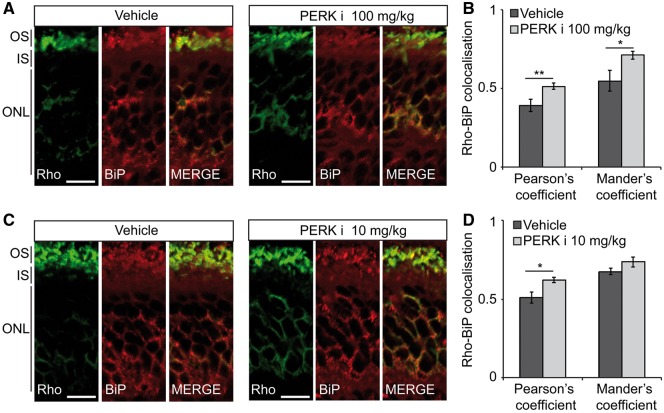
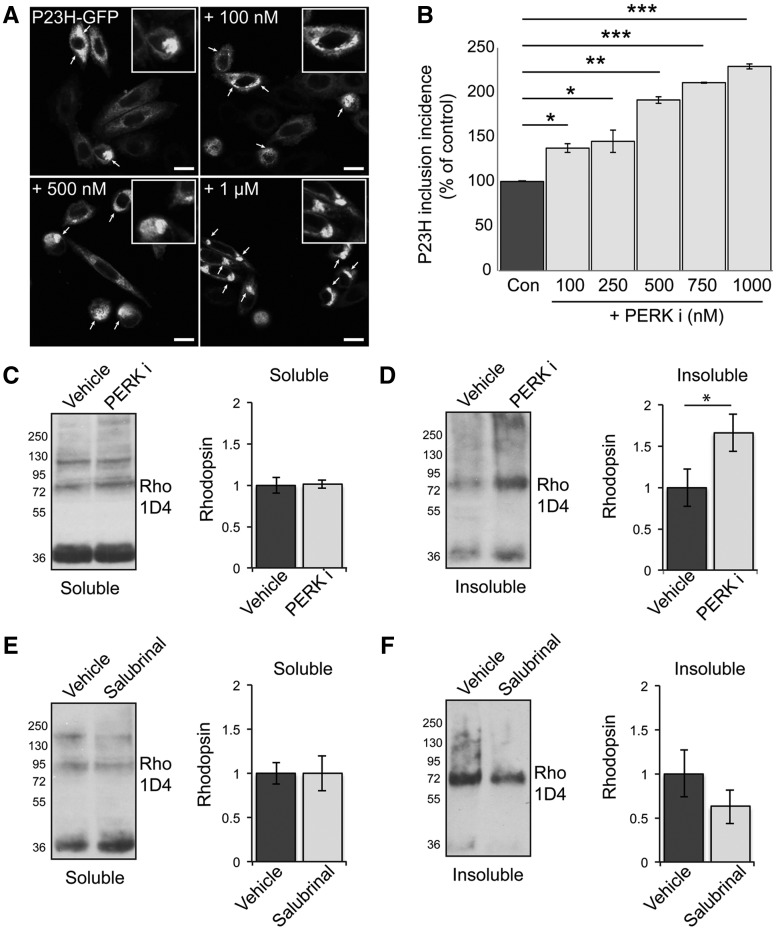
Mutations in rhodopsin, the light-sensitive protein of rod cells, are the most common cause of dominant retinitis pigmentosa (RP), a type of inherited blindness caused by the dysfunction and death of photoreceptor cells. The P23H mutation, the most frequent single cause of RP in the USA, causes rhodopsin misfolding and induction of the unfolded protein response (UPR), an adaptive ER stress response and signalling network that aims to enhance the folding and degradation of misfolded proteins to restore proteostasis. Prolonged UPR activation, and in particular the PERK branch, can reduce protein synthesis and initiate cell death through induction of pro-apoptotic pathways. Here, we investigated the effect of pharmacological PERK inhibition on retinal disease process in the P23H-1 transgenic rat model of retinal degeneration. PERK inhibition with GSK2606414A led to an inhibition of eIF2α phosphorylation, which correlated with reduced ERG function and decreased photoreceptor survival at both high and low doses of PERK inhibitor. Additionally, PERK inhibition increased the incidence of inclusion formation in cultured cells overexpressing P23H rod opsin, and increased rhodopsin aggregation in the P23H-1 rat retina, suggesting enhanced P23H misfolding and aggregation. In contrast, treatment of P23H-1 rats with an inhibitor of eIF2α phosphatase, salubrinal, led to improved photoreceptor survival. Collectively, these data suggest the activation of PERK is part of a protective response to mutant rhodopsin that ultimately limits photoreceptor cell death.
© The Author 2017. Published by Oxford University Press.
Figures
References
-
- Dryja T.P., McGee T.L., Reichel E., Hahn L.B., Cowley G.S., Yandell D.W., Sandberg M.A., Berson E.L. (1990) A point mutation of the rhodopsin gene in one form of retinitis pigmentosa. Nature, 343, 364–366. - PubMed
-
- Mendes H.F., van der Spuy J., Chapple J.P., Cheetham M.E. (2005) Mechanisms of cell death in rhodopsin retinitis pigmentosa: implications for therapy. Trends Mol. Med., 11, 177–185. - PubMed
-
- Illing M.E., Rajan R.S., Bence N.F., Kopito R.R. (2002) A rhodopsin mutant linked to autosomal dominant retinitis pigmentosa is prone to aggregate and interacts with the ubiquitin proteasome system. J. Biol. Chem., 277, 34150–34160. - PubMed
-
- Saliba R.S., Munro P.M., Luthert P.J., Cheetham M.E. (2002) The cellular fate of mutant rhodopsin: quality control, degradation and aggresome formation. J. Cell Sci., 115, 2907–2918. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
