

Genetic reduction of Nrf2 exacerbates cognitive deficits in a mouse model of Alzheimer's disease
- PMID: 29036636
- PMCID: PMC5886066
- DOI: 10.1093/hmg/ddx361
Genetic reduction of Nrf2 exacerbates cognitive deficits in a mouse model of Alzheimer's disease
Abstract
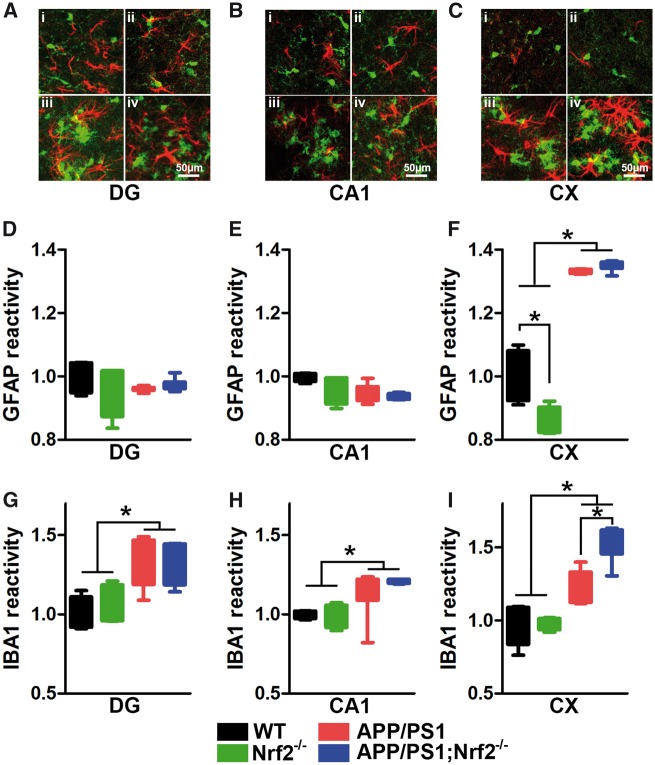
Aging is the major risk factor for several neurodegenerative diseases, including Alzheimer's disease (AD). However, the mechanisms by which aging contributes to neurodegeneration remain elusive. The nuclear factor (erythroid-derived 2)-like 2 (Nrf2) is a transcription factor that regulates expression of a vast number of genes by binding to the antioxidant response element. Nrf2 levels decrease as a function of age, and reduced Nrf2 levels have been reported in postmortem human brains and animal models of AD. Nevertheless, it is still unknown whether Nrf2 plays a role in the cognitive deficits associated with AD. To address this question, we used a genetic approach to remove the Nrf2 gene from APP/PS1 mice, a widely used animal model of AD. We found that the lack of Nrf2 significantly exacerbates cognitive deficits in APP/PS1, without altering gross motor function. Specifically, we found an exacerbation of deficits in spatial learning and memory, as well as in working and associative memory. Different brain regions control these behavioral tests, indicating that the lack of Nrf2 has a global effect on brain function. The changes in cognition were linked to an increase in Aβ and interferon-gamma (IFNγ) levels, and microgliosis. The changes in IFNγ levels are noteworthy as previously published evidence indicates that IFNγ can increase microglia activation and induce Aβ production. Our data suggest a clear link between Nrf2 and AD-mediated cognitive decline and further strengthen the connection between Nrf2 and AD.
© The Author 2017. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures
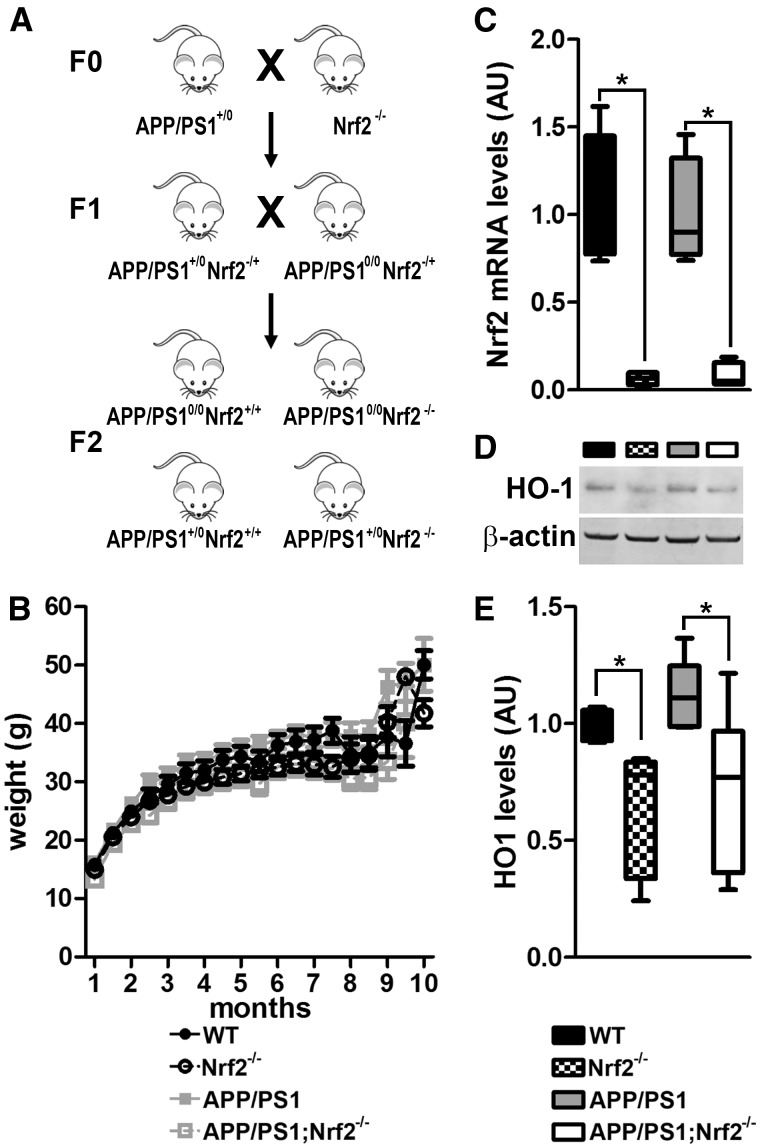
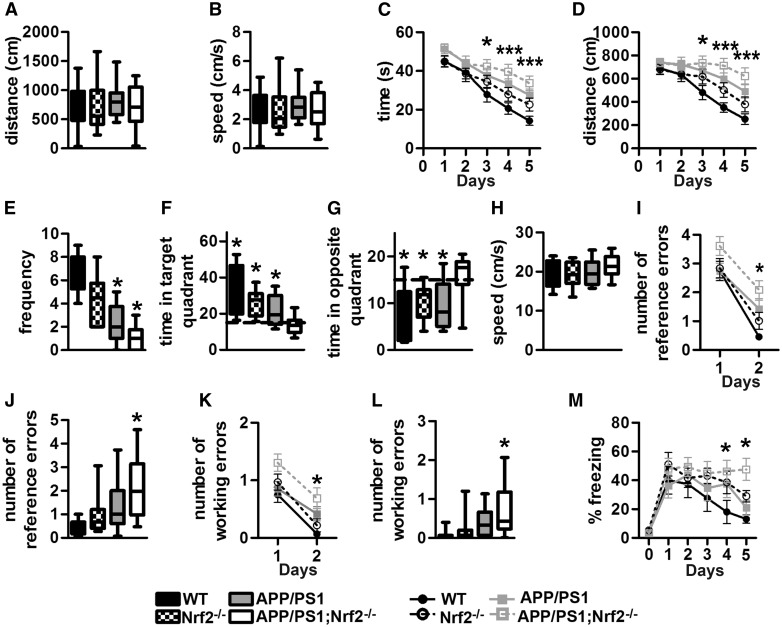
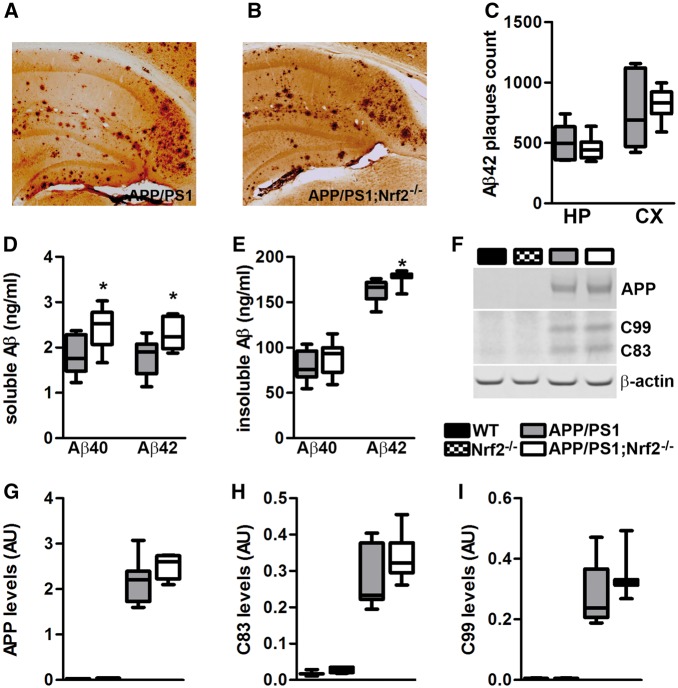
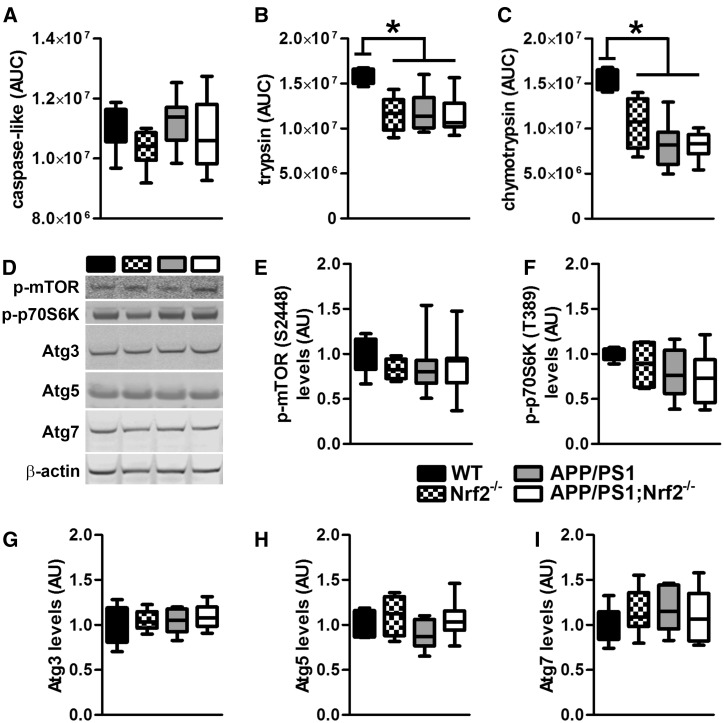

References
-
- Hayes J.D., Dinkova-Kostova A.T. (2014) The Nrf2 regulatory network provides an interface between redox and intermediary metabolism. Trends Biochem. Sci., 39, 199–218. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
