

Modulators of Enterococcus faecalis Cell Envelope Integrity and Antimicrobial Resistance Influence Stable Colonization of the Mammalian Gastrointestinal Tract
- PMID: 29038125
- PMCID: PMC5736811
- DOI: 10.1128/IAI.00381-17
Modulators of Enterococcus faecalis Cell Envelope Integrity and Antimicrobial Resistance Influence Stable Colonization of the Mammalian Gastrointestinal Tract
Abstract
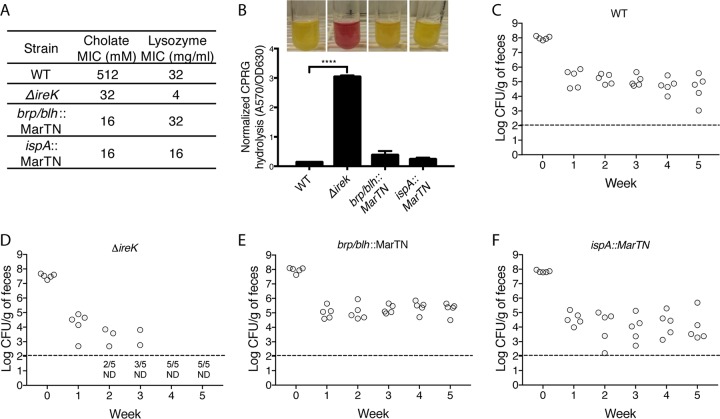
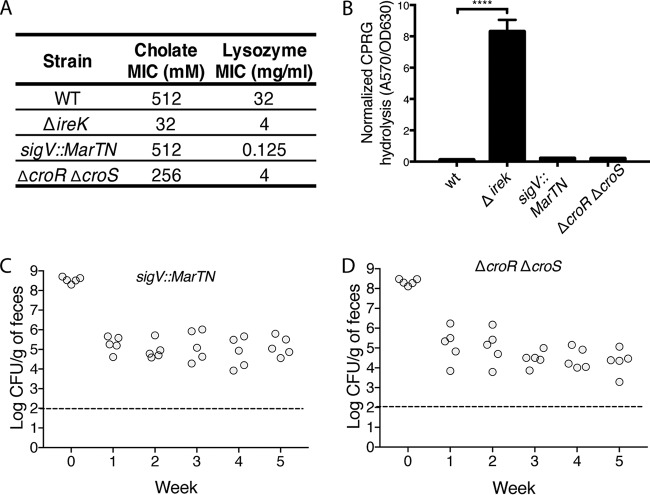
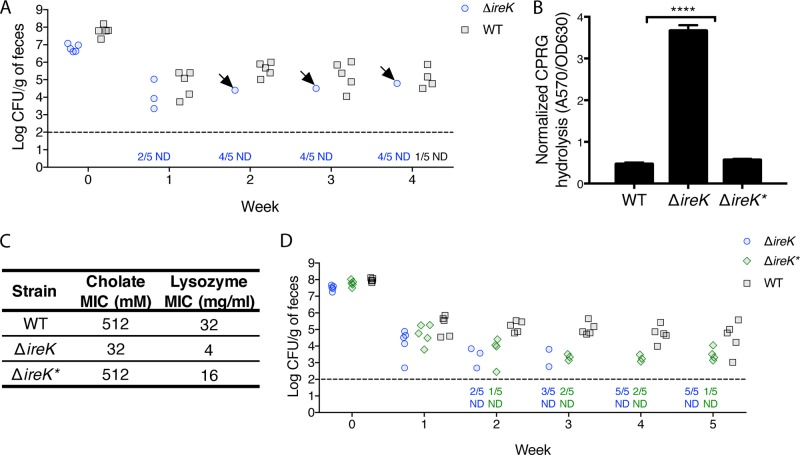
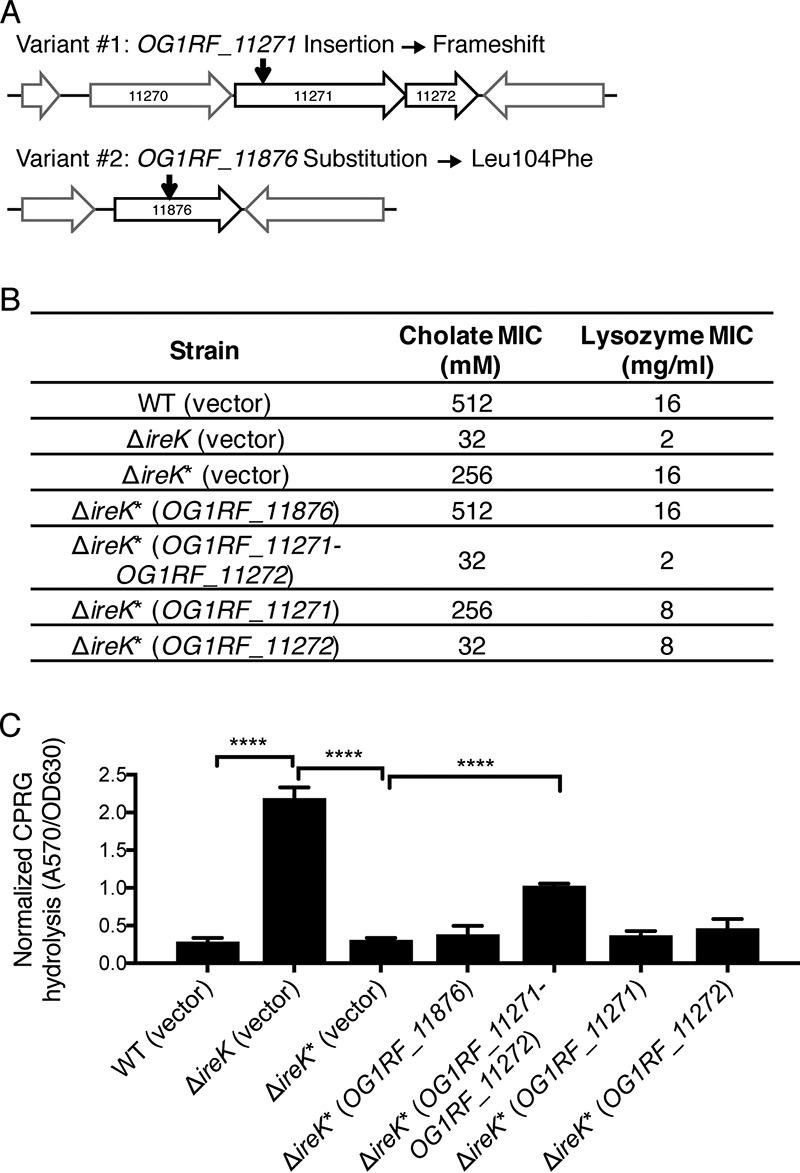
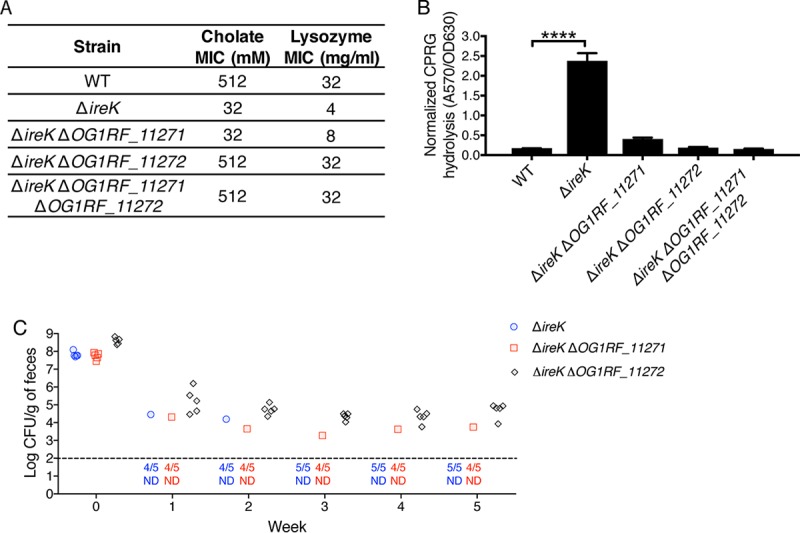
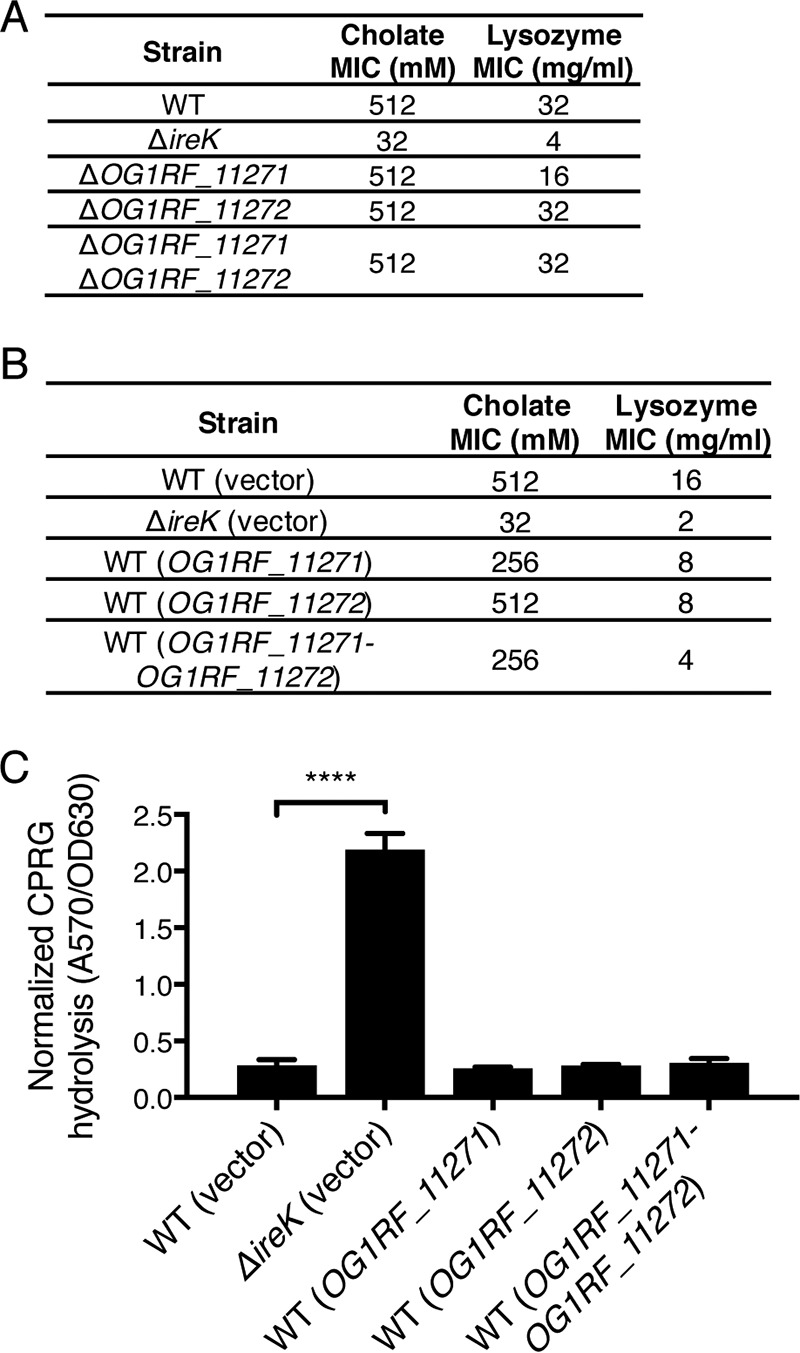
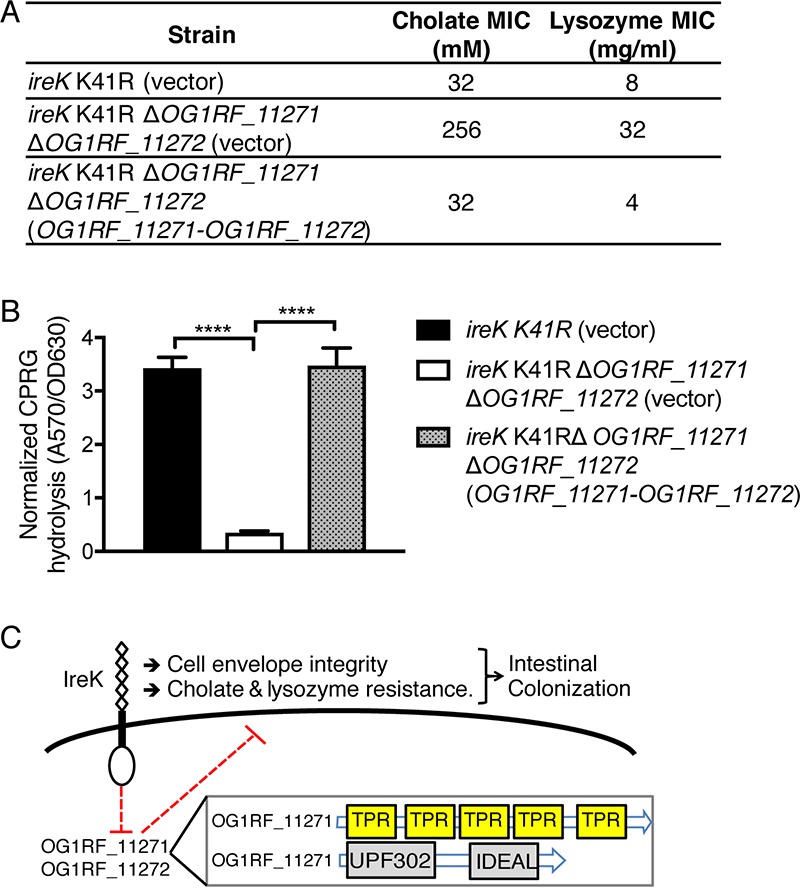
The Gram-positive bacterium Enterococcus faecalis is both a colonizer of the gastrointestinal tract (GIT) and an agent of serious nosocomial infections. Although it is typically required for pathogenesis, GIT colonization by E. faecalis is poorly understood. E. faecalis tolerates high concentrations of GIT antimicrobials, like cholate and lysozyme, leading us to hypothesize that resistance to intestinal antimicrobials is essential for long-term GIT colonization. Analyses of E. faecalis mutants exhibiting defects in antimicrobial resistance revealed that IreK, a determinant of envelope integrity and antimicrobial resistance, is required for long-term GIT colonization. IreK is a member of the PASTA kinase protein family, bacterial transmembrane signaling proteins implicated in the regulation of cell wall homeostasis. Among several determinants of cholate and lysozyme resistance in E. faecalis, IreK was the only one found to be required for intestinal colonization, emphasizing the importance of this protein to enterococcal adaptation to the GIT. By studying ΔireK suppressor mutants that recovered the ability to colonize the GIT, we identified two conserved enterococcal proteins (OG1RF_11271 and OG1RF_11272) that function antagonistically to IreK and interfere with cell envelope integrity, antimicrobial resistance, and GIT colonization. Our data suggest that IreK, through its kinase activity, inhibits the actions of these proteins. IreK, OG1RF_11271, and OG1RF_11272 are found in all enterococci, suggesting that their effect on GIT colonization is universal across enterococci. Thus, we have defined conserved genes in the enterococcal core genome that influence GIT colonization through their effect on enterococcal envelope integrity and antimicrobial resistance.
Keywords: Enterococcus; IreK; antimicrobial resistance; cell envelope integrity; colonization.
Copyright © 2017 American Society for Microbiology.
Figures
Similar articles
-
Convergence of PASTA Kinase and Two-Component Signaling in Response to Cell Wall Stress in Enterococcus faecalis.J Bacteriol. 2018 May 24;200(12):e00086-18. doi: 10.1128/JB.00086-18. Print 2018 Jun 15. J Bacteriol. 2018. PMID: 29632091 Free PMC article.
-
IreK-Mediated, Cell Wall-Protective Phosphorylation in Enterococcus faecalis.J Proteome Res. 2021 Nov 5;20(11):5131-5144. doi: 10.1021/acs.jproteome.1c00635. Epub 2021 Oct 21. J Proteome Res. 2021. PMID: 34672600 Free PMC article.
-
Reciprocal regulation of cephalosporin resistance in Enterococcus faecalis.mBio. 2011 Nov 1;2(6):e00199-11. doi: 10.1128/mBio.00199-11. Print 2011. mBio. 2011. PMID: 22045988 Free PMC article.
-
The enterococcal PASTA kinase: A sentinel for cell envelope stress.Mol Oral Microbiol. 2021 Apr;36(2):132-144. doi: 10.1111/omi.12313. Epub 2020 Oct 5. Mol Oral Microbiol. 2021. PMID: 32945615 Free PMC article. Review.
-
Colonization of the mammalian intestinal tract by enterococci.Curr Opin Microbiol. 2019 Feb;47:26-31. doi: 10.1016/j.mib.2018.10.005. Epub 2018 Nov 13. Curr Opin Microbiol. 2019. PMID: 30439685 Free PMC article. Review.
Cited by
-
Insights into ecology, pathogenesis, and biofilm formation of Enterococcus faecalis from functional genomics.Microbiol Mol Biol Rev. 2025 Mar 27;89(1):e0008123. doi: 10.1128/mmbr.00081-23. Epub 2024 Dec 23. Microbiol Mol Biol Rev. 2025. PMID: 39714182 Review.
-
Urinary Tract Infection and Antimicrobial Susceptibility of Bacterial Isolates in Saint Joseph Kitgum Hospital, Kitgum, Uganda.Antibiotics (Basel). 2022 Apr 11;11(4):504. doi: 10.3390/antibiotics11040504. Antibiotics (Basel). 2022. PMID: 35453255 Free PMC article.
-
Reciprocal Regulation of PASTA Kinase Signaling by Differential Modification.J Bacteriol. 2019 Apr 24;201(10):e00016-19. doi: 10.1128/JB.00016-19. Print 2019 May 15. J Bacteriol. 2019. PMID: 30858297 Free PMC article.
-
Characterization of an Enterococcus faecalis bacteriophage SFQ1 as a potential therapeutic agent.Front Microbiol. 2023 Jun 22;14:1210319. doi: 10.3389/fmicb.2023.1210319. eCollection 2023. Front Microbiol. 2023. PMID: 37426023 Free PMC article.
-
Disruption of the tagF Orthologue in the epa Locus Variable Region of Enterococcus faecalis Causes Cell Surface Changes and Suppresses an eep-Dependent Lysozyme Resistance Phenotype.J Bacteriol. 2022 Oct 18;204(10):e0024722. doi: 10.1128/jb.00247-22. Epub 2022 Sep 12. J Bacteriol. 2022. PMID: 36094307 Free PMC article.
References
-
- Donskey CJ, Chowdhry TK, Hecker MT, Hoyen CK, Hanrahan JA, Hujer AM, Hutton-Thomas RA, Whalen CC, Bonomo RA, Rice LB. 2000. Effect of antibiotic therapy on the density of vancomycin-resistant enterococci in the stool of colonized patients. N Engl J Med 343:1925–1932. doi:10.1056/NEJM200012283432604. - DOI - PMC - PubMed
-
- Ubeda C, Taur Y, Jenq RR, Equinda MJ, Son T, Samstein M, Viale A, Socci ND, van den Brink MRM, Kamboj M, Pamer EG. 2010. Vancomycin-resistant Enterococcus domination of intestinal microbiota is enabled by antibiotic treatment in mice and precedes bloodstream invasion in humans. J Clin Invest 120:4332–4341. doi:10.1172/JCI43918. - DOI - PMC - PubMed
-
- Top J, Paganelli FL, Zhang X, van Schaik W, Leavis HL, van Luit-Asbroek M, van der Poll T, Leendertse M, Bonten MJ, Willems RJ. 2013. The Enterococcus faecium enterococcal biofilm regulator, EbrB, regulates the esp operon and is implicated in biofilm formation and intestinal colonization. PLoS One 8:e65224. doi:10.1371/journal.pone.0065224. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
