

Studies of Pseudomonas aeruginosa Mutants Indicate Pyoverdine as the Central Factor in Inhibition of Aspergillus fumigatus Biofilm
- PMID: 29038255
- PMCID: PMC5717155
- DOI: 10.1128/JB.00345-17
Studies of Pseudomonas aeruginosa Mutants Indicate Pyoverdine as the Central Factor in Inhibition of Aspergillus fumigatus Biofilm
Abstract
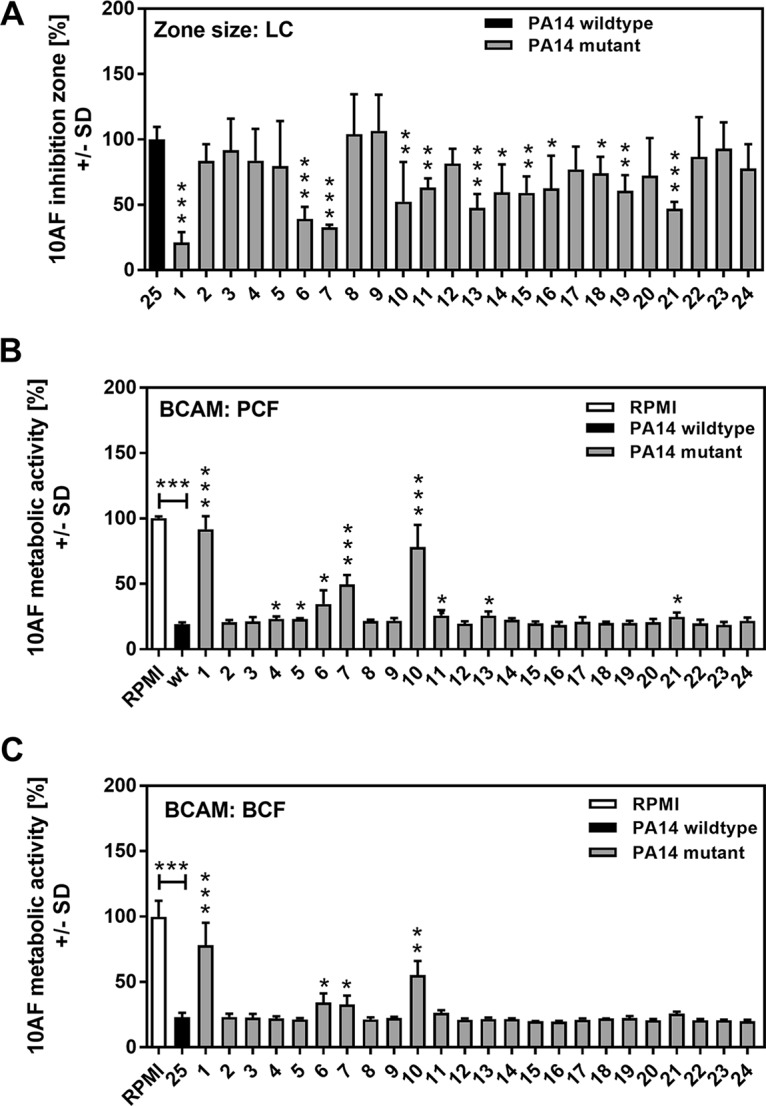
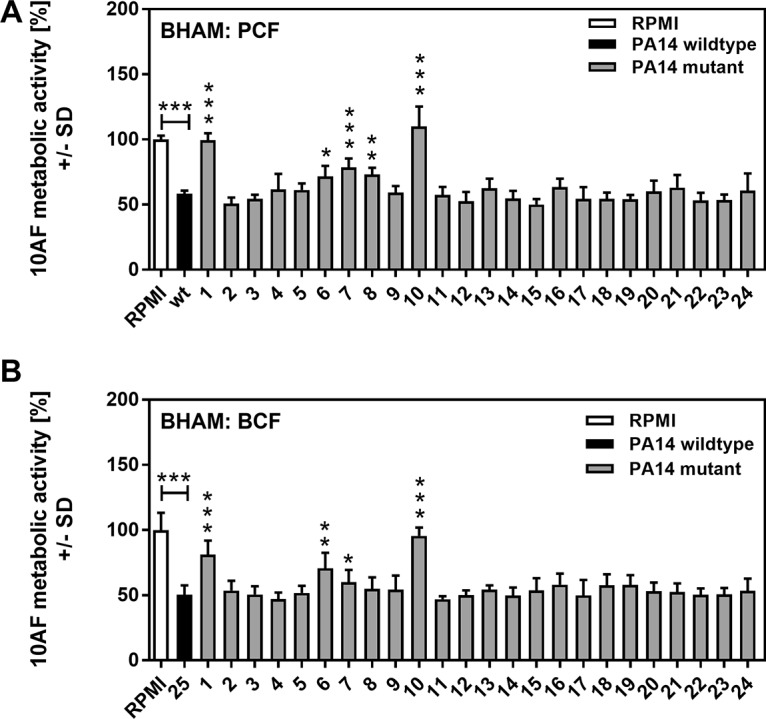
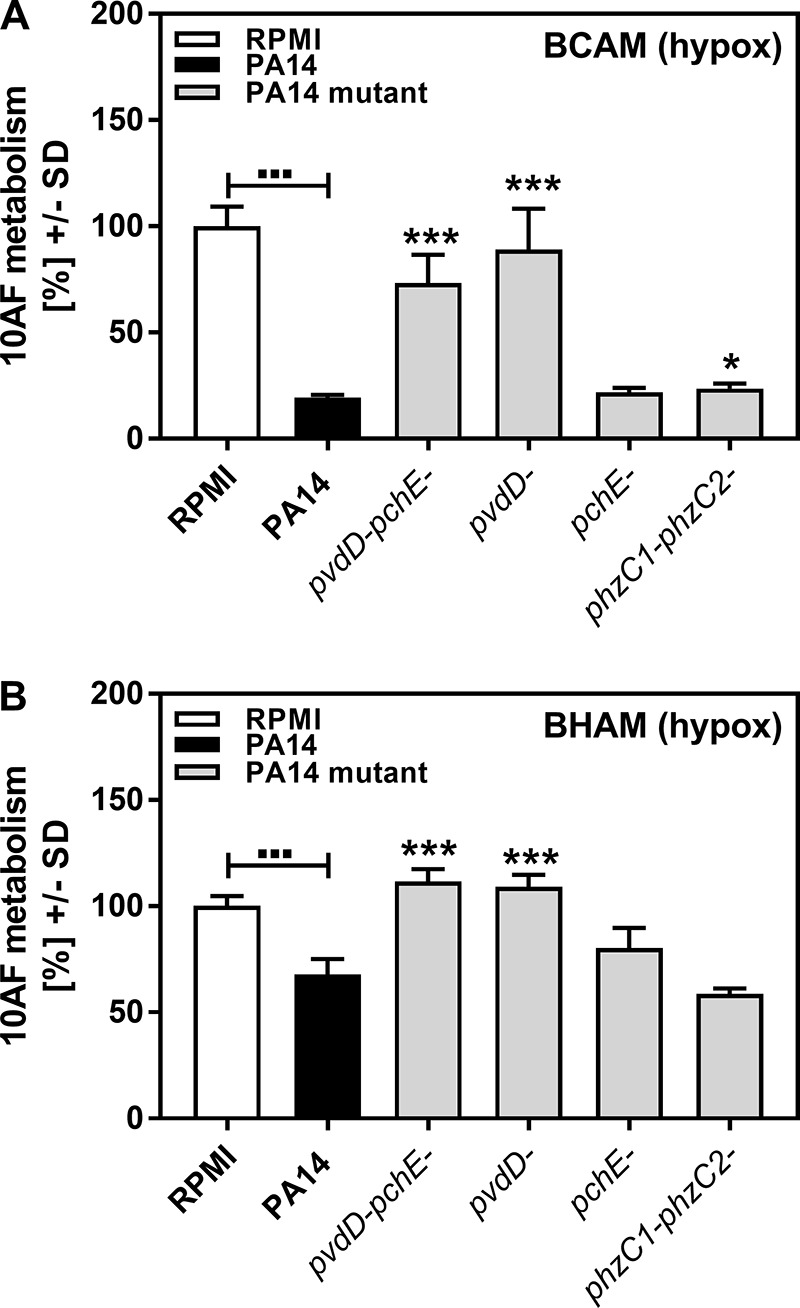
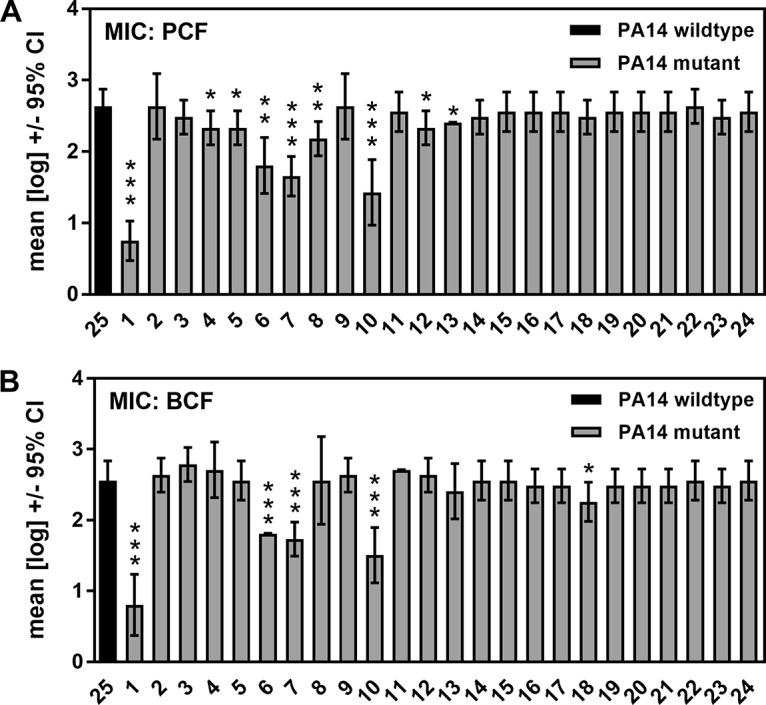
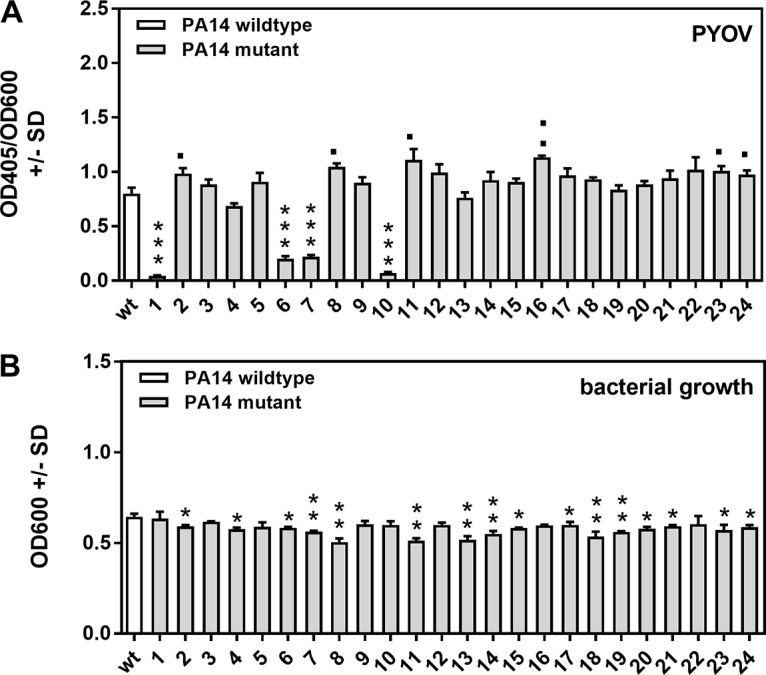
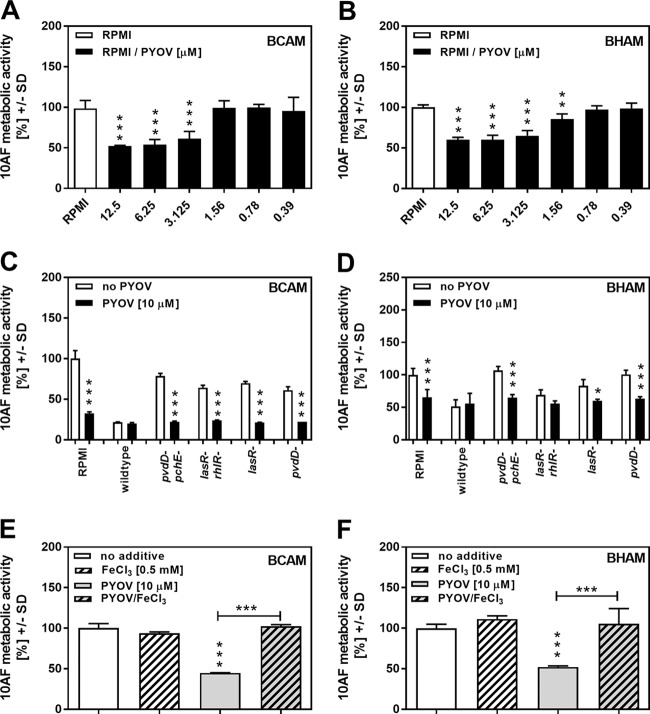
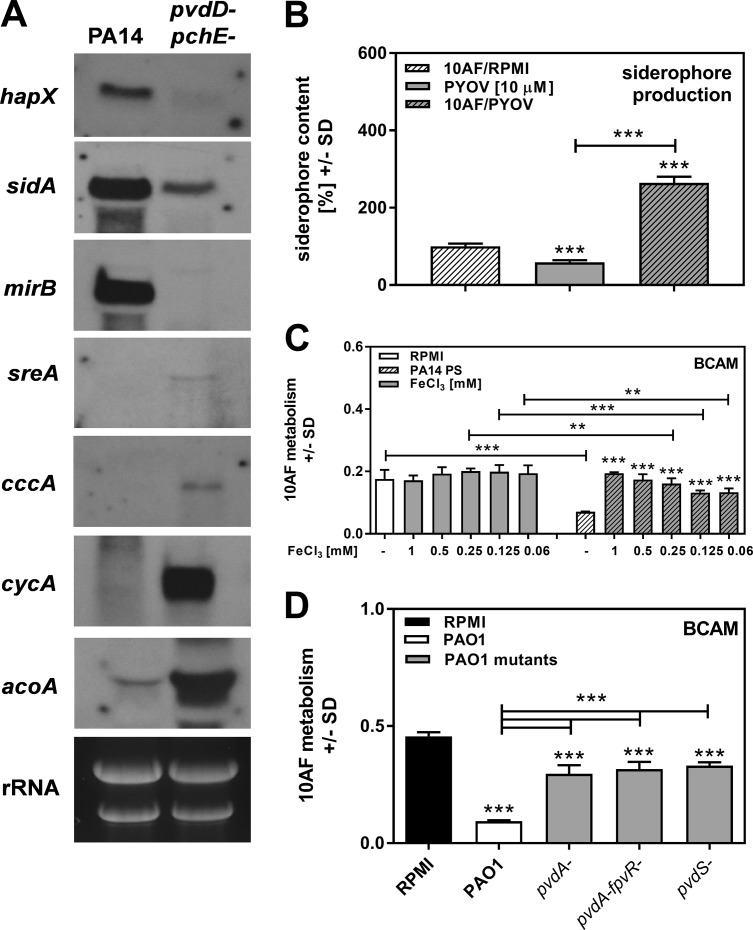
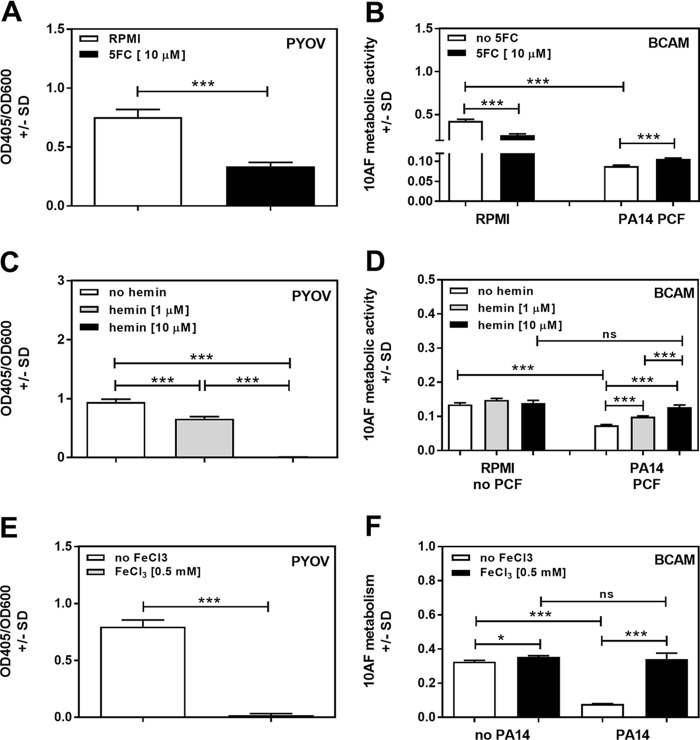
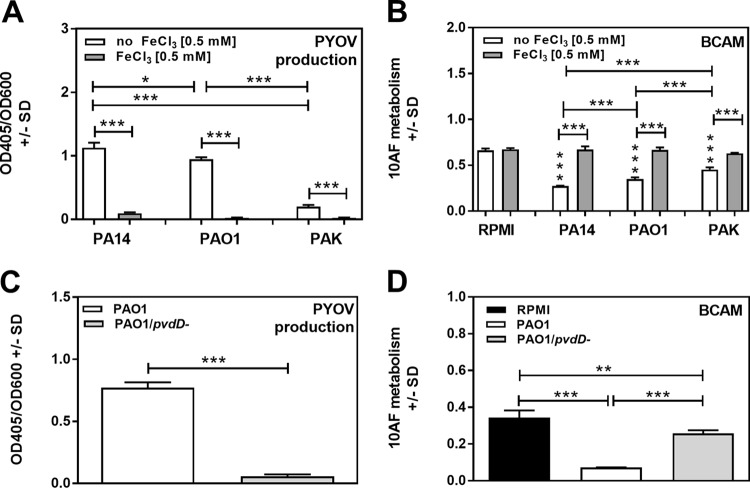
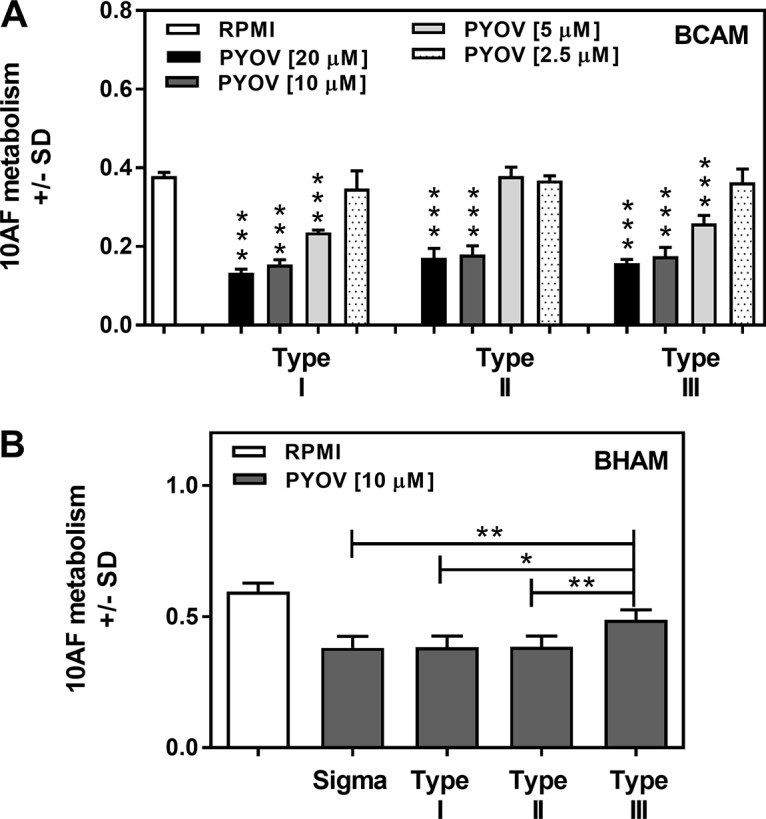
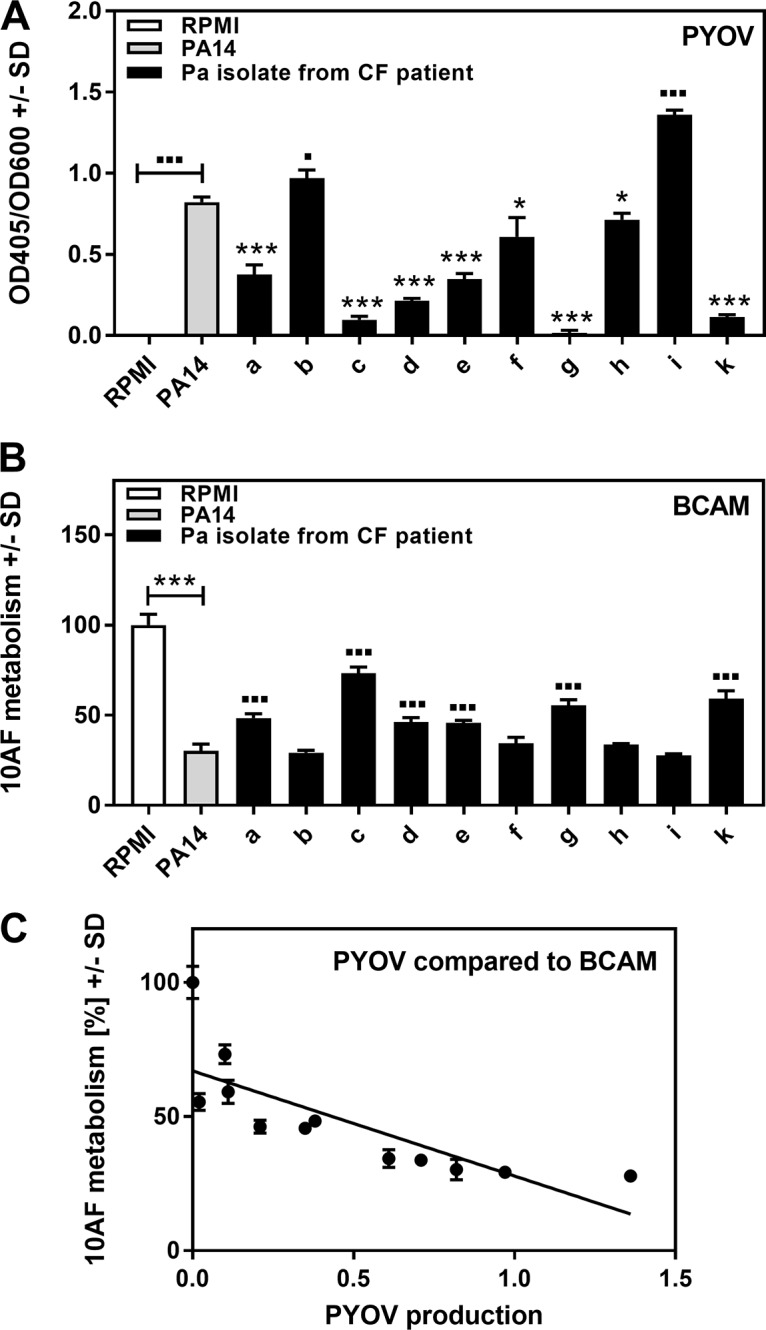
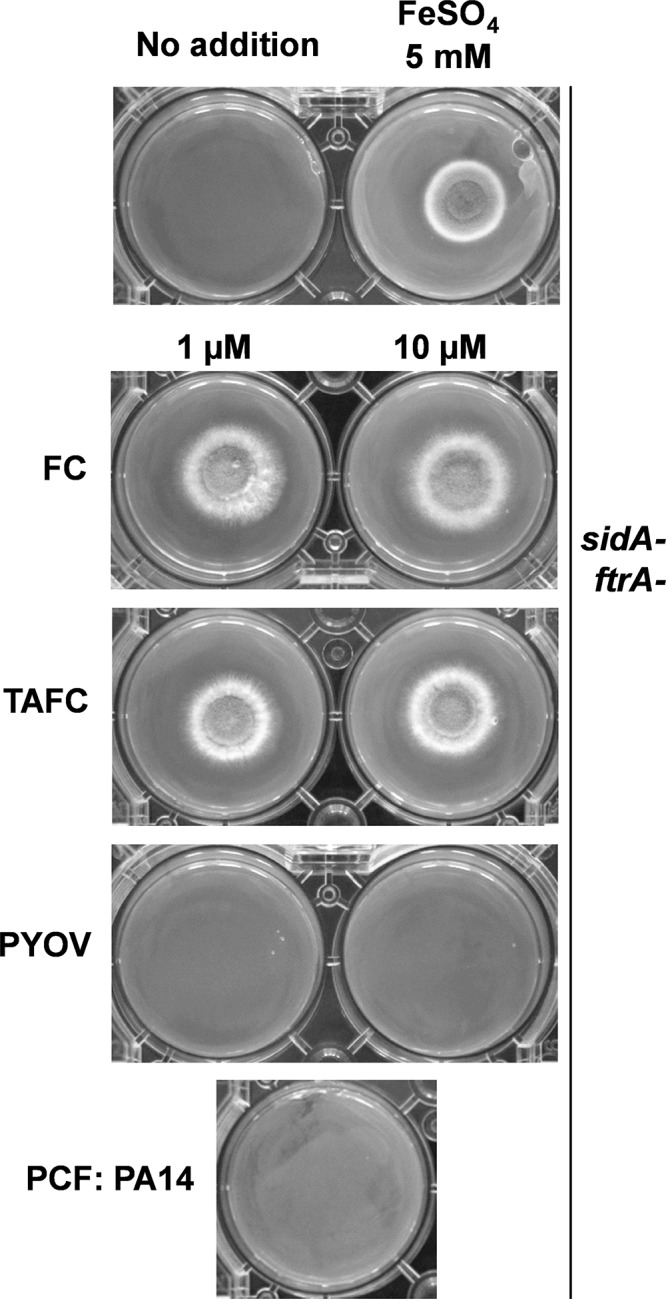
Pseudomonas aeruginosa and Aspergillus fumigatus are common opportunistic bacterial and fungal pathogens, respectively. They often coexist in airways of immunocompromised patients and individuals with cystic fibrosis, where they form biofilms and cause acute and chronic illnesses. Hence, the interactions between them have long been of interest and it is known that P. aeruginosa can inhibit A. fumigatusin vitro We have approached the definition of the inhibitory P. aeruginosa molecules by studying 24 P. aeruginosa mutants with various virulence genes deleted for the ability to inhibit A. fumigatus biofilms. The ability of P. aeruginosa cells or their extracellular products produced during planktonic or biofilm growth to affect A. fumigatus biofilm metabolism or planktonic A. fumigatus growth was studied in agar and liquid assays using conidia or hyphae. Four mutants, the pvdD pchE, pvdD, lasR rhlR, and lasR mutants, were shown to be defective in various assays. This suggested the P. aeruginosa siderophore pyoverdine as the key inhibitory molecule, although additional quorum sensing-regulated factors likely contribute to the deficiency of the latter two mutants. Studies of pure pyoverdine substantiated these conclusions and included the restoration of inhibition by the pyoverdine deletion mutants. A correlation between the concentration of pyoverdine produced and antifungal activity was also observed in clinical P. aeruginosa isolates derived from lungs of cystic fibrosis patients. The key inhibitory mechanism of pyoverdine was chelation of iron and denial of iron to A. fumigatusIMPORTANCE Interactions between human pathogens found in the same body locale are of vast interest. These interactions could result in exacerbation or amelioration of diseases. The bacterium Pseudomonas aeruginosa affects the growth of the fungus Aspergillus fumigatus Both pathogens form biofilms that are resistant to therapeutic drugs and host immunity. P. aeruginosa and A. fumigatus biofilms are found in vivo, e.g., in the lungs of cystic fibrosis patients. Studying 24 P. aeruginosa mutants, we identified pyoverdine as the major anti-A. fumigatus compound produced by P. aeruginosa Pyoverdine captures iron from the environment, thus depriving A. fumigatus of a nutrient essential for its growth and metabolism. We show how microbes of different kingdoms compete for essential resources. Iron deprivation could be a therapeutic approach to the control of pathogen growth.
Keywords: Aspergillus fumigatus; Pseudomonas aeruginosa; biofilms; intermicrobial interaction; iron; mutants; pyoverdine.
Copyright © 2017 American Society for Microbiology.
Figures
Similar articles
-
Aspergillus-Pseudomonas interaction, relevant to competition in airways.Med Mycol. 2019 Apr 1;57(Supplement_2):S228-S232. doi: 10.1093/mmy/myy087. Med Mycol. 2019. PMID: 30816973 Review.
-
Intermicrobial interaction: Aspergillus fumigatus siderophores protect against competition by Pseudomonas aeruginosa.PLoS One. 2019 May 8;14(5):e0216085. doi: 10.1371/journal.pone.0216085. eCollection 2019. PLoS One. 2019. PMID: 31067259 Free PMC article.
-
Under nonlimiting iron conditions pyocyanin is a major antifungal molecule, and differences between prototypic Pseudomonas aeruginosa strains.Med Mycol. 2021 May 4;59(5):453-464. doi: 10.1093/mmy/myaa066. Med Mycol. 2021. PMID: 32827431
-
Small Colony Variants of Pseudomonas aeruginosa Display Heterogeneity in Inhibiting Aspergillus fumigatus Biofilm.Mycopathologia. 2018 Feb;183(1):263-272. doi: 10.1007/s11046-017-0186-9. Epub 2017 Aug 7. Mycopathologia. 2018. PMID: 28785939
-
Pseudomonas aeruginosa virulence attenuation by inhibiting siderophore functions.Appl Microbiol Biotechnol. 2023 Feb;107(4):1019-1038. doi: 10.1007/s00253-022-12347-6. Epub 2023 Jan 12. Appl Microbiol Biotechnol. 2023. PMID: 36633626 Review.
Cited by
-
The Expanding Mycovirome of Aspergilli.J Fungi (Basel). 2024 Aug 17;10(8):585. doi: 10.3390/jof10080585. J Fungi (Basel). 2024. PMID: 39194910 Free PMC article. Review.
-
Co-Operative Biofilm Interactions between Aspergillus fumigatus and Pseudomonas aeruginosa through Secreted Galactosaminogalactan Exopolysaccharide.J Fungi (Basel). 2022 Mar 24;8(4):336. doi: 10.3390/jof8040336. J Fungi (Basel). 2022. PMID: 35448567 Free PMC article.
-
Synthesis of the Hydroxamate Siderophore Nα-Methylcoprogen B in Scedosporium apiospermum Is Mediated by sidD Ortholog and Is Required for Virulence.Front Cell Infect Microbiol. 2020 Oct 28;10:587909. doi: 10.3389/fcimb.2020.587909. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 33194829 Free PMC article.
-
Same Game, Different Players: Emerging Pathogens of the CF Lung.mBio. 2021 Jan 12;12(1):e01217-20. doi: 10.1128/mBio.01217-20. mBio. 2021. PMID: 33436426 Free PMC article. Review.
-
Methodologies for in vitro and in vivo evaluation of efficacy of antifungal and antibiofilm agents and surface coatings against fungal biofilms.Microb Cell. 2018 Jun 14;5(7):300-326. doi: 10.15698/mic2018.07.638. Microb Cell. 2018. PMID: 29992128 Free PMC article. Review.
References
-
- Smyth AR, Hurley MN. 2010. Targeting the Pseudomonas aeruginosa biofilm to combat infections in patients with cystic fibrosis. Drugs Future 35:1007–1014. doi:10.1358/dof.2010.035.012.1537937. - DOI
-
- Fillaux J, Brémont F, Murris M, Cassaing S, Rittié JL, Tétu L, Segonds C, Abbal M, Bieth E, Berry A, Pipy B, Magnaval JF. 2012. Assessment of Aspergillus sensitization or persistent carriage as a factor in lung function impairment in cystic fibrosis patients. Scand J Infect Dis 44:842–847. doi:10.3109/00365548.2012.695454. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
