

The small GTPase RSG1 controls a final step in primary cilia initiation
- PMID: 29038301
- PMCID: PMC5748968
- DOI: 10.1083/jcb.201604048
The small GTPase RSG1 controls a final step in primary cilia initiation
Abstract
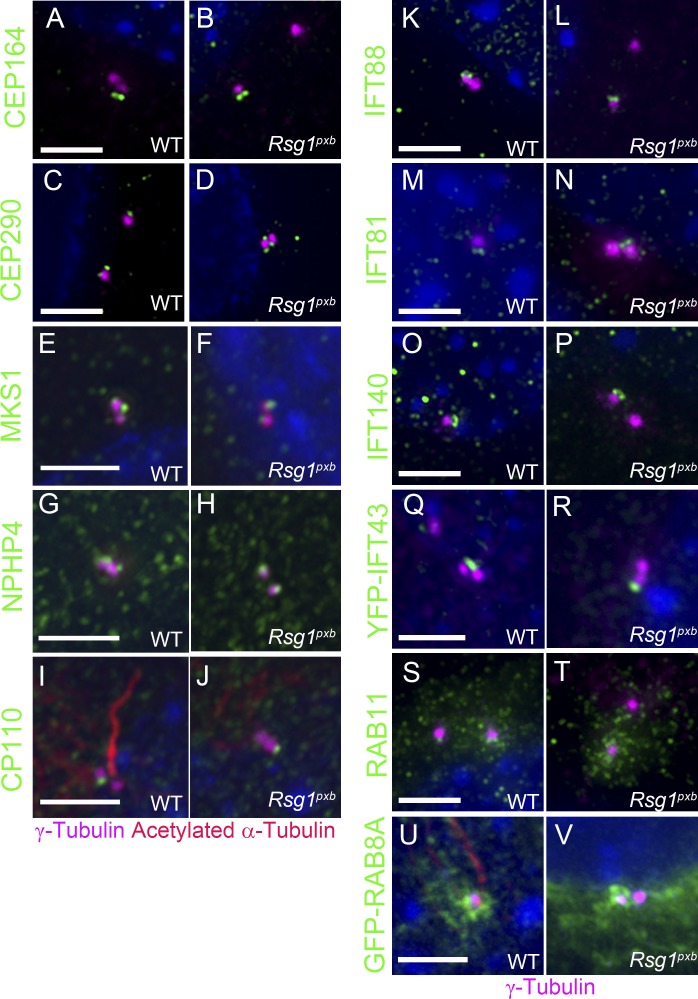
Primary cilia, which are essential for normal development and tissue homeostasis, are extensions of the mother centriole, but the mechanisms that remodel the centriole to promote cilia initiation are poorly understood. Here we show that mouse embryos that lack the small guanosine triphosphatase RSG1 die at embryonic day 12.5, with developmental abnormalities characteristic of decreased cilia-dependent Hedgehog signaling. Rsg1 mutant embryos have fewer primary cilia than wild-type embryos, but the cilia that form are of normal length and traffic Hedgehog pathway proteins within the cilium correctly. Rsg1 mother centrioles recruit proteins required for cilia initiation and dock onto ciliary vesicles, but axonemal microtubules fail to elongate normally. RSG1 localizes to the mother centriole in a process that depends on tau tubulin kinase 2 (TTBK2), the CPLANE complex protein Inturned (INTU), and its own GTPase activity. The data suggest a specific role for RSG1 in the final maturation of the mother centriole and ciliary vesicle that allows extension of the ciliary axoneme.
© 2018 Agbu et al.
Figures







References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
