

Outer inflammatory protein a (OipA) of Helicobacter pylori is regulated by host cell contact and mediates CagA translocation and interleukin-8 response only in the presence of a functional cag pathogenicity island type IV secretion system
- PMID: 29040466
- PMCID: PMC6433299
- DOI: 10.1093/femspd/ftx113
Outer inflammatory protein a (OipA) of Helicobacter pylori is regulated by host cell contact and mediates CagA translocation and interleukin-8 response only in the presence of a functional cag pathogenicity island type IV secretion system
Abstract
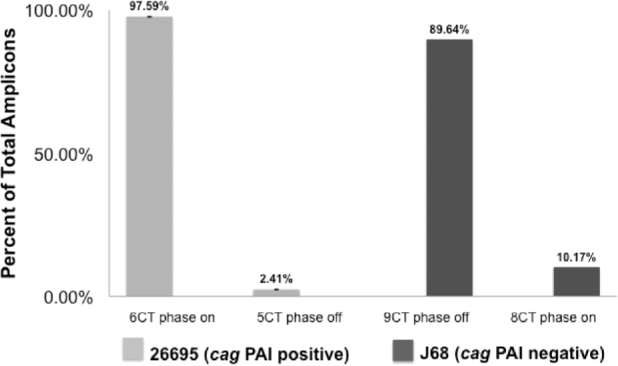
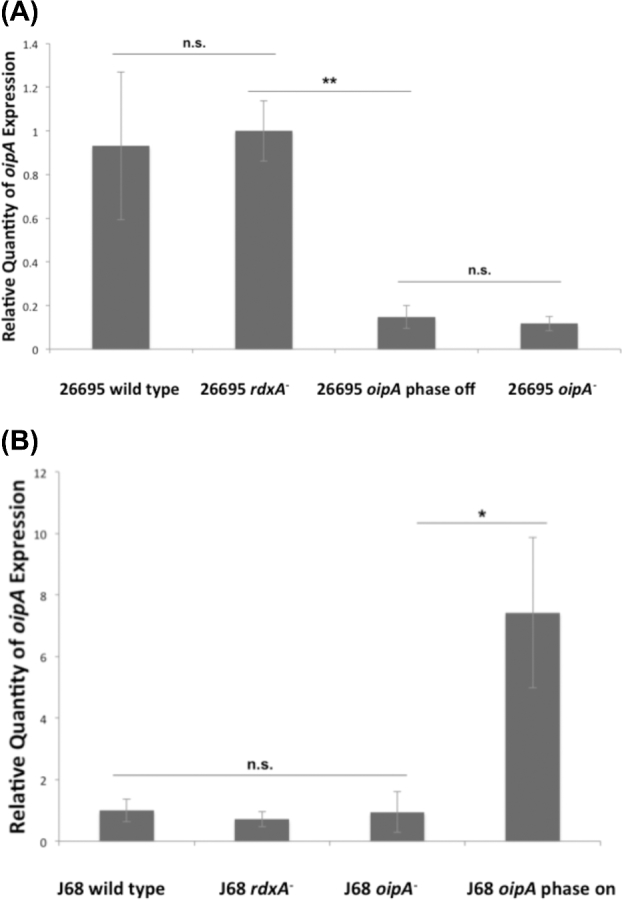
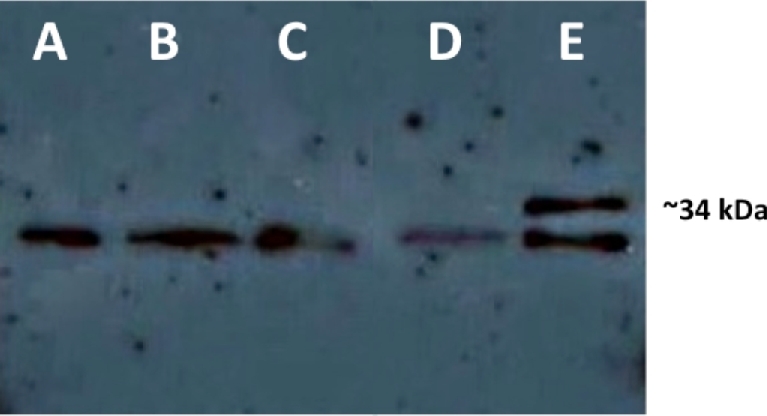
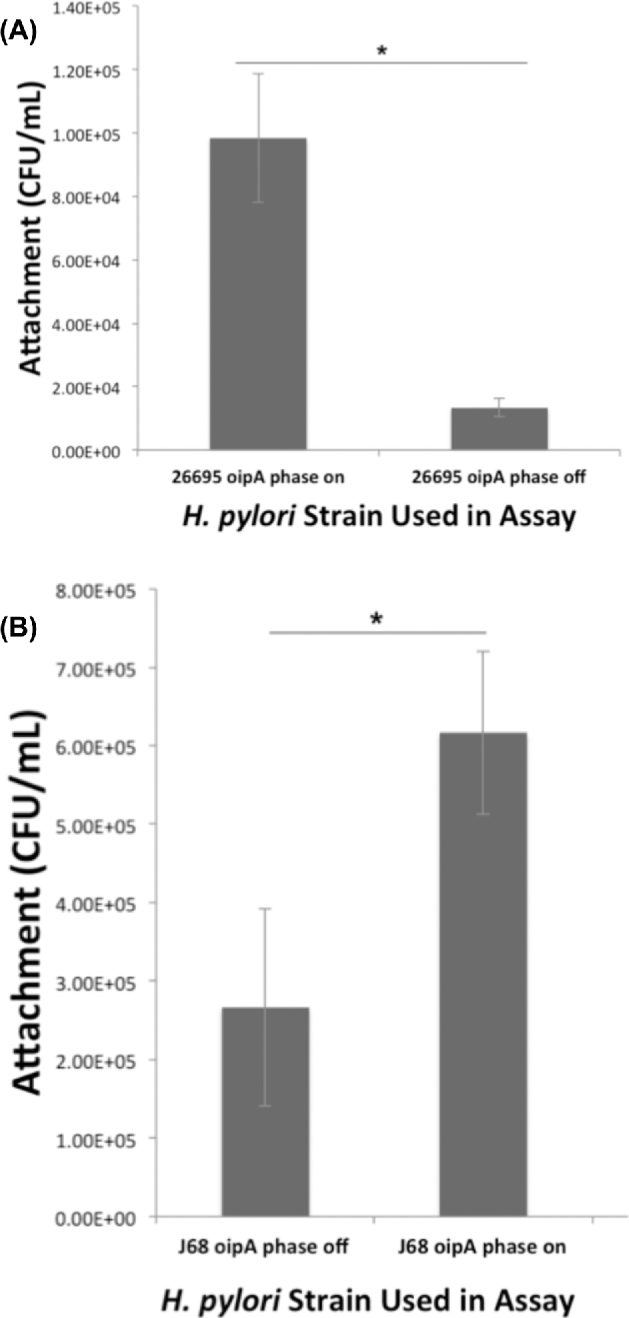
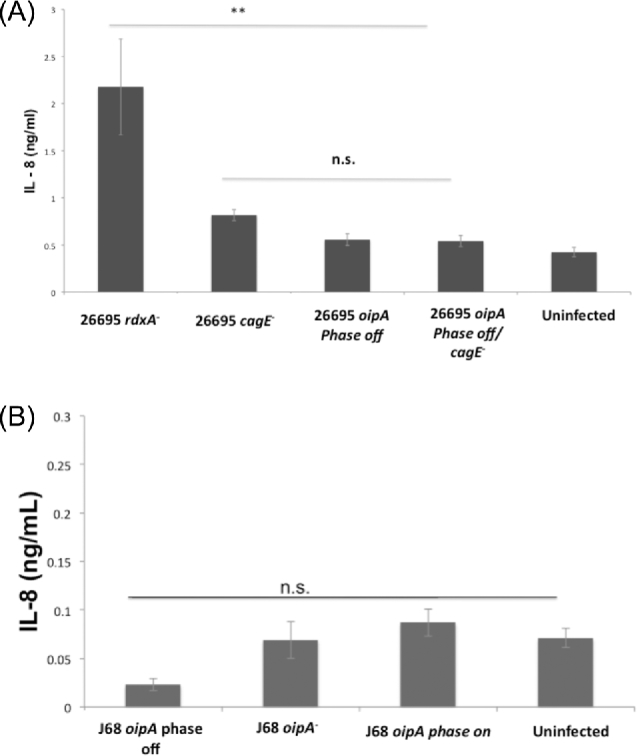
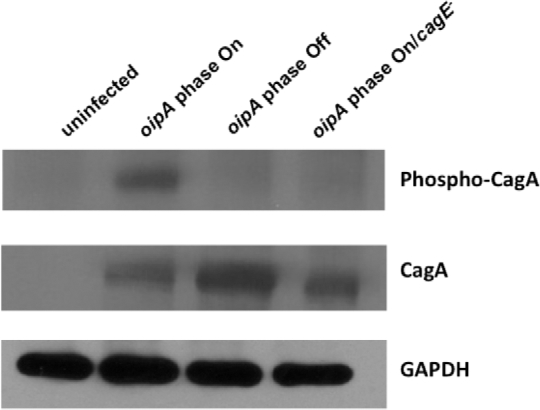
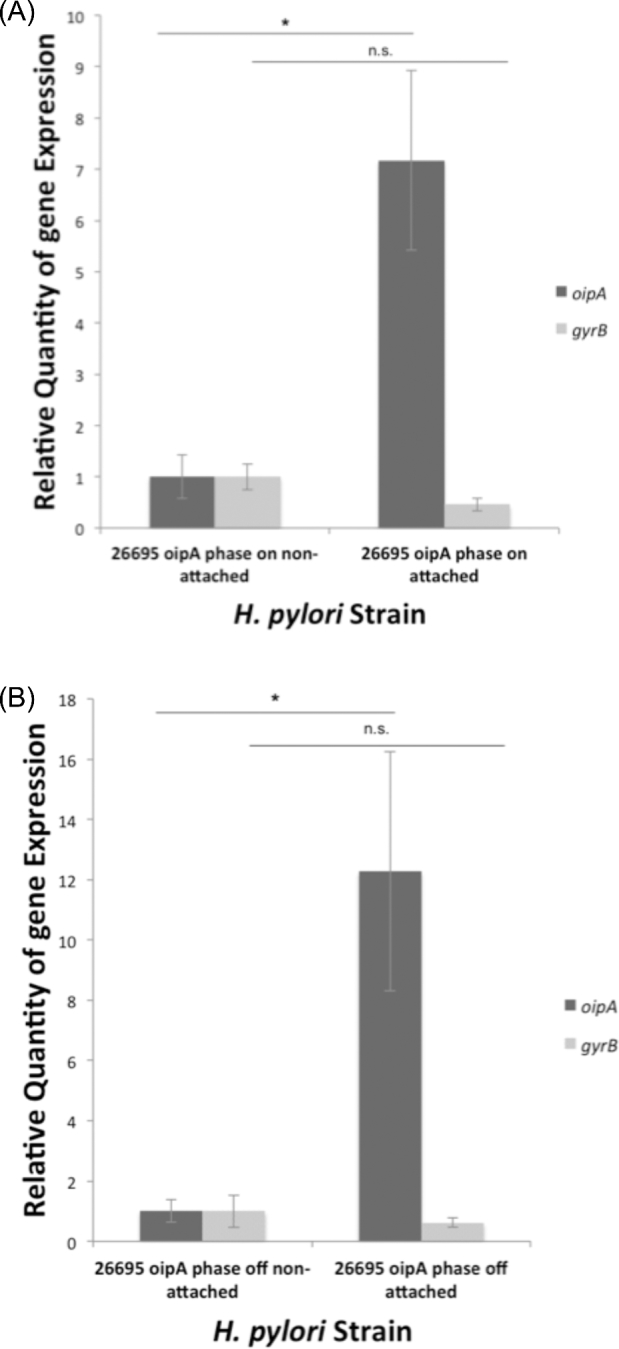
OipA is a phase-variable virulence factor of Helicobacter pylori. Mutations in oipA to turn the gene phase on in a cag pathogenicity island (PAI)-negative strain of H. pylori (J68) or phase off in a cag PAI-positive strain (26695) demonstrated that phase on oipA alleles in both strains had both increased oipA mRNA and human gastric adenocarcinoma (AGS) cell adherence compared to isogenic oipA phase off mutants. An oipA phase off mutant of H. pylori 26695 demonstrated decreased IL-8 secretion by AGS cells and failure to translocate the cag PAI effector CagA. Increased attachment by OipA expressing cag PAI-negative H. pylori J68 failed to alter secreted IL-8 levels. Thus, OipA is necessary but not sufficient for the induction of IL-8; however, it is necessary for translocation of the oncoprotein CagA. Perhaps the nearly invariant phase on status of oipA alleles among cag PAI-positive H. pylori isolates relates to the role of this outer membrane protein in effective translocation of CagA. oipA mRNA comparisons between AGS cell-adherent and non-adherent H. pylori 26695 revealed significantly greater levels in the adherent cells. This may allow H. pylori to adapt to conditions of host cell contact by altering expression of this virulence factor.
Keywords: CagA translocation; adhesin; interleukin-8; phase variation.
© FEMS 2017. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
References
-
- Acio-Pizzarello CR, Acio AA, Choi EJ et al. Determinants of the regulation of H. pylori adhesins include repeat sequences in both promoter and coding regions as well as the two component system, ArsRS. J Med Microbiol 2017;66:798–807. - PubMed
-
- Chen J, Lin M, Li N et al. Therapeutic vaccination with Salmonella-derived codon optimized outer inflammatory protein DNA vaccine enhances protection in Helicobacter pylori infected mice. Vaccine 2012;30:5310–5. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
