

Mutual maintenance of di- and triploid Pelophylax esculentus hybrids in R-E systems: results from artificial crossings experiments
- PMID: 29041900
- PMCID: PMC5645918
- DOI: 10.1186/s12862-017-1063-3
Mutual maintenance of di- and triploid Pelophylax esculentus hybrids in R-E systems: results from artificial crossings experiments
Abstract
Background: Interspecies animal hybrids can employ clonal or hemiclonal reproduction modes where one or all parental genomes are transmitted to the progeny without recombination. Nevertheless, some interspecies hybrids retain strong connection with the parental species needed for successful reproduction. Appearance of polyploid hybrid animals may play an important role in the substitution of parental species and in the speciation process.
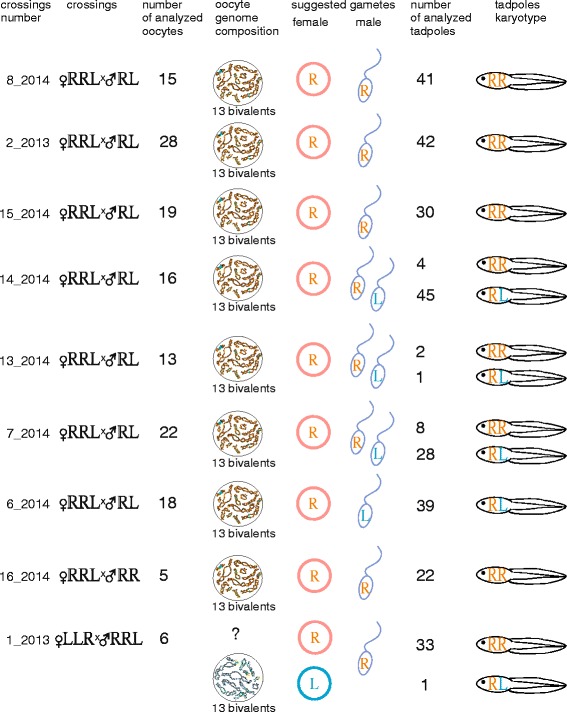
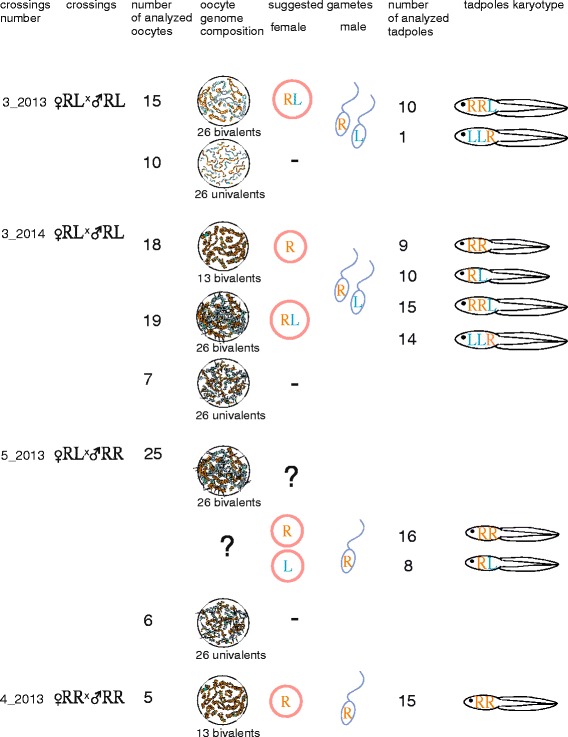
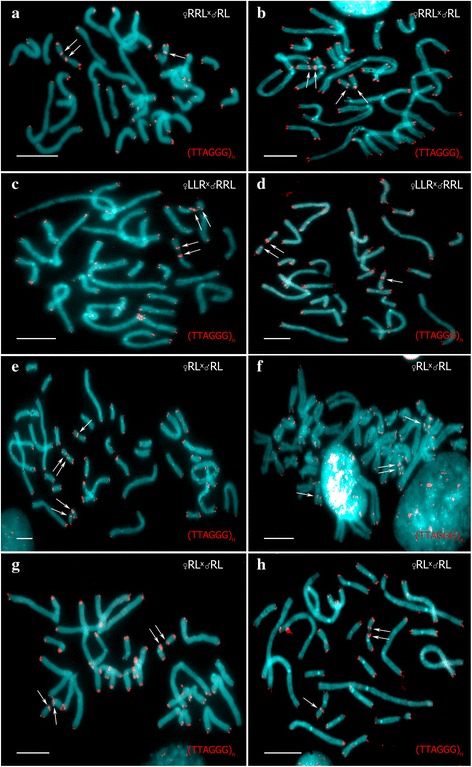
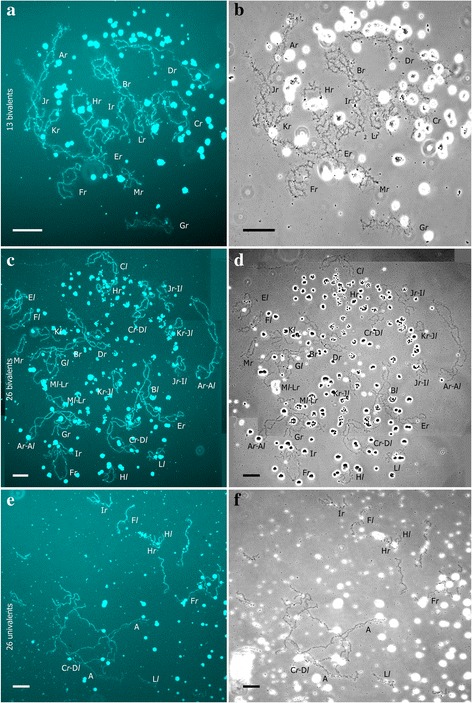
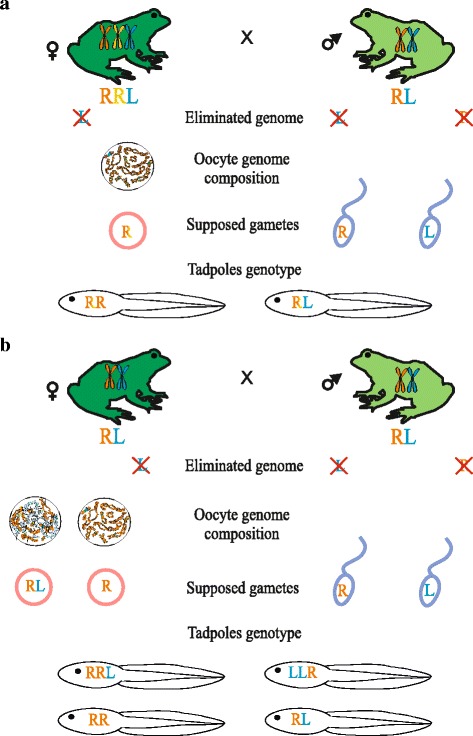
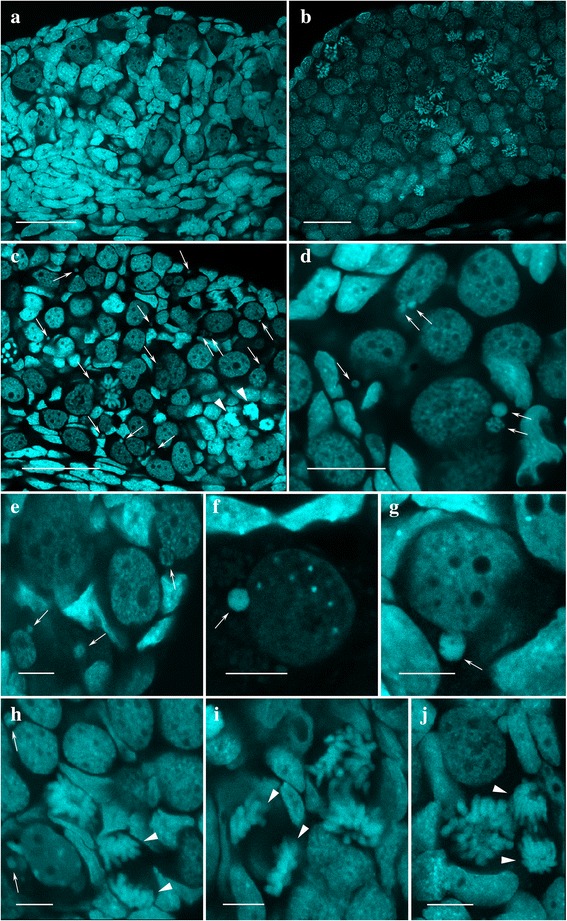
Results: To establish the mechanisms that enable parental species, diploid and polyploid hybrids coexist we have performed artificial crossing experiments of water frogs of Pelophylax esculentus complex. We identified tadpole karyotypes and oocyte genome composition in all females involved in the crossings. The majority of diploid and triploid hybrid frogs produced oocytes with 13 bivalents leading to haploid gametes with the same genome as parental species hybrids usually coexist with. After fertilization of such gametes only diploid animals appeared. Oocytes with 26 bivalents produced by some diploid hybrid frogs lead to diploid gametes, which give rise to triploid hybrids after fertilization. In gonads of all diploid and triploid hybrid tadpoles we found DAPI-positive micronuclei (nucleus-like bodies) involved in selective genome elimination. Hybrid male and female individuals produced tadpoles with variable karyotype and ploidy even in one crossing owing to gametes with various genome composition.
Conclusions: We propose a model of diploid and triploid hybrid frog reproduction in R-E population systems. Triploid Pelophylax esculentus hybrids can transmit genome of parental species they coexist with by producing haploid gametes with the same genome composition. Triploid hybrids cannot produce triploid individuals after crossings with each other and depend on diploid hybrid females producing diploid eggs. In contrast to other population systems, the majority of diploid and triploid hybrid females unexpectedly produced gametes with the same genome as parental species hybrids coexist with.
Keywords: Gamete; Genome elimination; Hemiclonal reproduction; Hybrid population systems; Karyotype; Polyploid hybrid.
Conflict of interest statement
Ethics approval and consent to participate
All manipulations with animals were carried out in accordance with the national and international guidelines. The field studies did not involve endangered or protected species. All specimens were collected in the regions of Ukraine, which are not considered as protected areas, thus no specific permissions were required for these locations. Techniques used to capture, breeding, tissue sampling and euthanasia sought to minimize animal suffering. Each individual was anaesthetized by methoxyethane or submersion in a 1% solution of 3-aminobenzoic acid ethyl ester (MS 222). All procedures were approved by the Local Animal Ethic Commettee of Saint-Petersburg State University (## 131–03-2 and 131–03-3).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
Similar articles
-
Maintenance of pure hybridogenetic water frog populations: Genotypic variability in progeny of diploid and triploid parents.PLoS One. 2022 Jul 6;17(7):e0268574. doi: 10.1371/journal.pone.0268574. eCollection 2022. PLoS One. 2022. PMID: 35793279 Free PMC article.
-
Variation in hybridogenetic hybrid emergence between populations of water frogs from the Pelophylax esculentus complex.PLoS One. 2019 Nov 1;14(11):e0224759. doi: 10.1371/journal.pone.0224759. eCollection 2019. PLoS One. 2019. PMID: 31675368 Free PMC article.
-
Gamete types, sex determination and stable equilibria of all-hybrid populations of diploid and triploid edible frogs (Pelophylax esculentus).BMC Evol Biol. 2009 Jun 15;9:135. doi: 10.1186/1471-2148-9-135. BMC Evol Biol. 2009. PMID: 19527499 Free PMC article.
-
Distant hybridization leads to different ploidy fishes.Sci China Life Sci. 2010 Apr;53(4):416-25. doi: 10.1007/s11427-010-0057-9. Epub 2010 May 7. Sci China Life Sci. 2010. PMID: 20596907 Review.
-
Genomic constitution and atypical reproduction in polyploid and unisexual lineages of the Misgurnus loach, a teleost fish.Cytogenet Genome Res. 2013;140(2-4):226-40. doi: 10.1159/000353301. Epub 2013 Jul 9. Cytogenet Genome Res. 2013. PMID: 23899809 Review.
Cited by
-
Effects of minimum Strigea robusta (Digenea: Strigeidae) cercariae doses and localization of cysts on the anomaly P manifestation in Pelophylax lessonae (Anura: Ranidae) tadpoles.Parasitol Res. 2023 Mar;122(3):889-894. doi: 10.1007/s00436-022-07778-z. Epub 2023 Jan 12. Parasitol Res. 2023. PMID: 36631683
-
Maintenance of pure hybridogenetic water frog populations: Genotypic variability in progeny of diploid and triploid parents.PLoS One. 2022 Jul 6;17(7):e0268574. doi: 10.1371/journal.pone.0268574. eCollection 2022. PLoS One. 2022. PMID: 35793279 Free PMC article.
-
Capture and return of sexual genomes by hybridogenetic frogs provide clonal genome enrichment in a sexual species.Sci Rep. 2021 Jan 15;11(1):1633. doi: 10.1038/s41598-021-81240-5. Sci Rep. 2021. PMID: 33452404 Free PMC article.
-
The programmed DNA elimination and formation of micronuclei in germ line cells of the natural hybridogenetic water frog Pelophylax esculentus.Sci Rep. 2018 May 18;8(1):7870. doi: 10.1038/s41598-018-26168-z. Sci Rep. 2018. PMID: 29777142 Free PMC article.
-
Variation in hybridogenetic hybrid emergence between populations of water frogs from the Pelophylax esculentus complex.PLoS One. 2019 Nov 1;14(11):e0224759. doi: 10.1371/journal.pone.0224759. eCollection 2019. PLoS One. 2019. PMID: 31675368 Free PMC article.
References
-
- Dawley RM, Bogart JP. Evolution and ecology of unisexual vertebrates. Albany, New York: New York State Museum Publications; 1989.
-
- Schön I, Martens K, van Dijk P. Lost sex. The evolutionary biology of parthenogenesis. Heidelberg, Germany: Springer; 2009.
-
- Vrijenhoek RC. Animal clones and diversity. Bioscience. 1998;48:617–628. doi: 10.2307/1313421. - DOI
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
