

Plant pathogenesis-related proteins of the cacao fungal pathogen Moniliophthora perniciosa differ in their lipid-binding specificities
- PMID: 29042440
- PMCID: PMC5733592
- DOI: 10.1074/jbc.M117.811398
Plant pathogenesis-related proteins of the cacao fungal pathogen Moniliophthora perniciosa differ in their lipid-binding specificities
Abstract
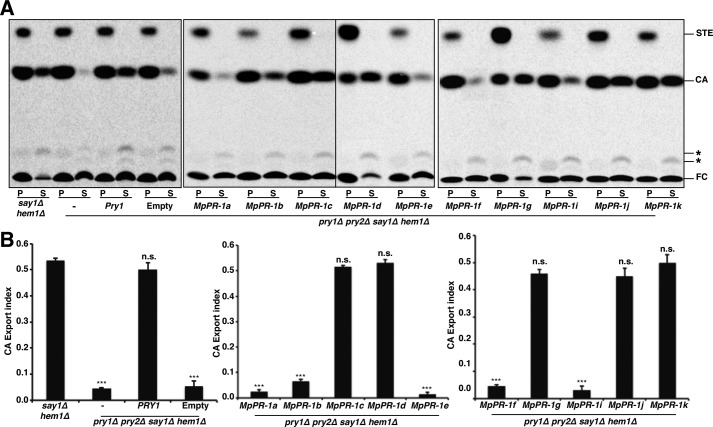
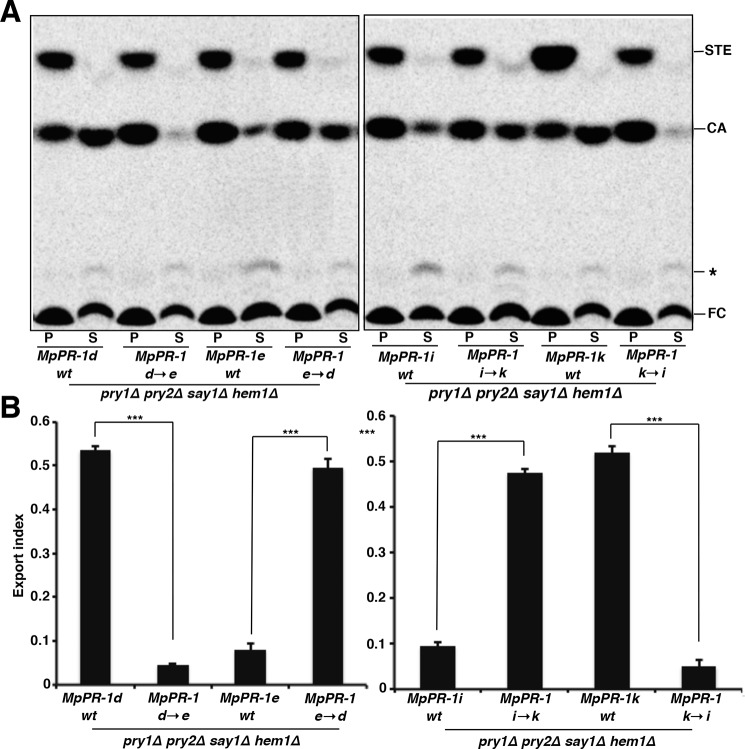
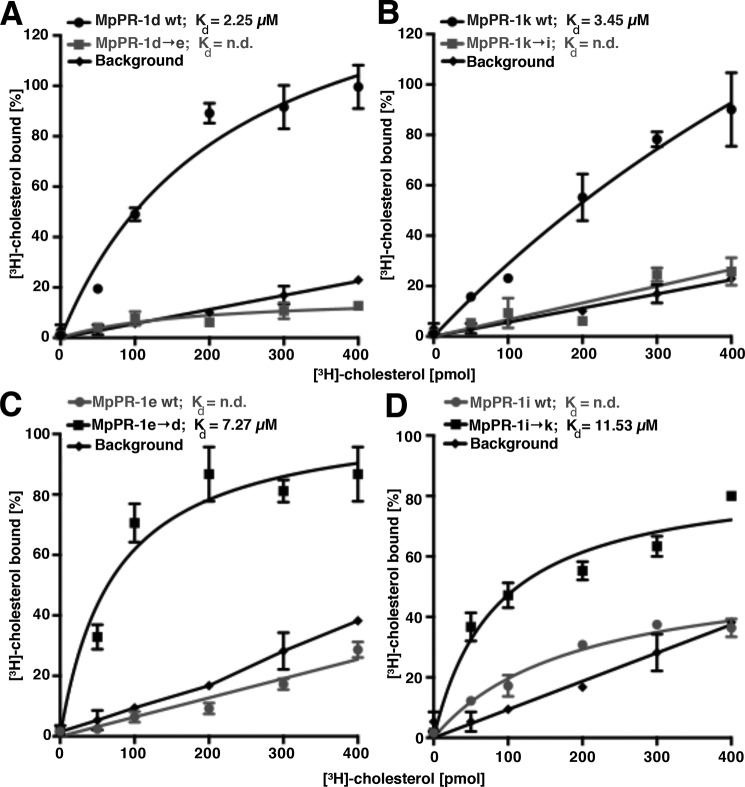
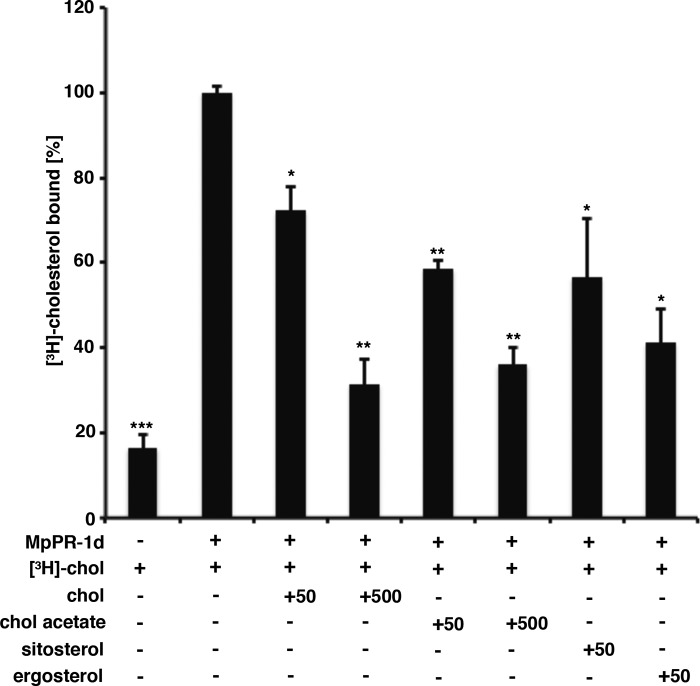
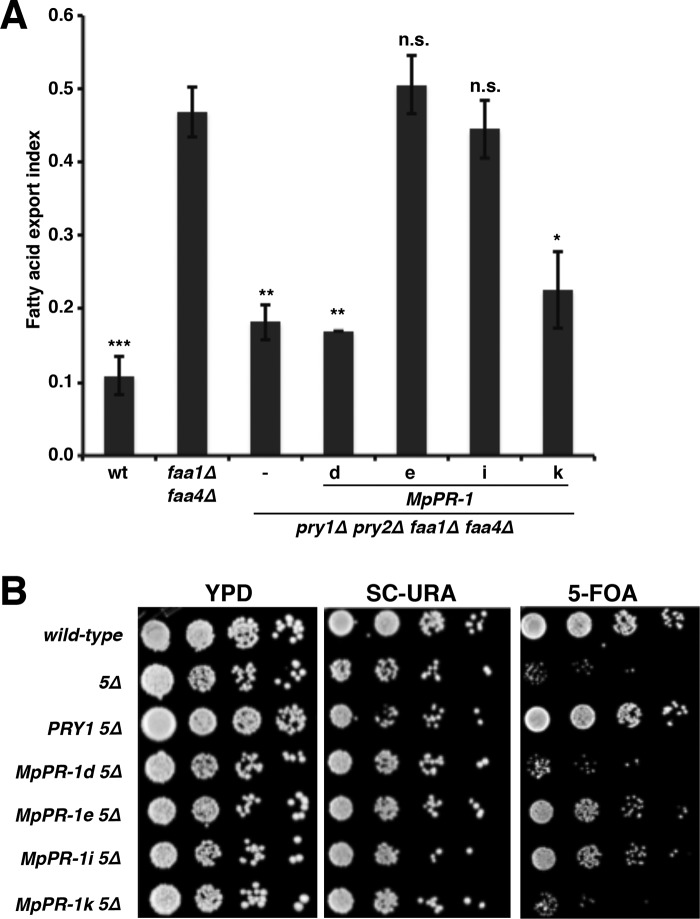
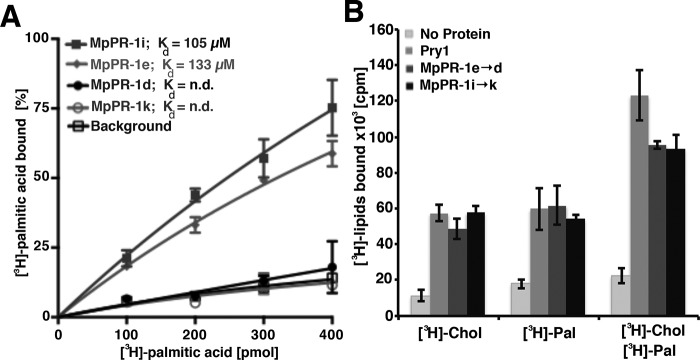
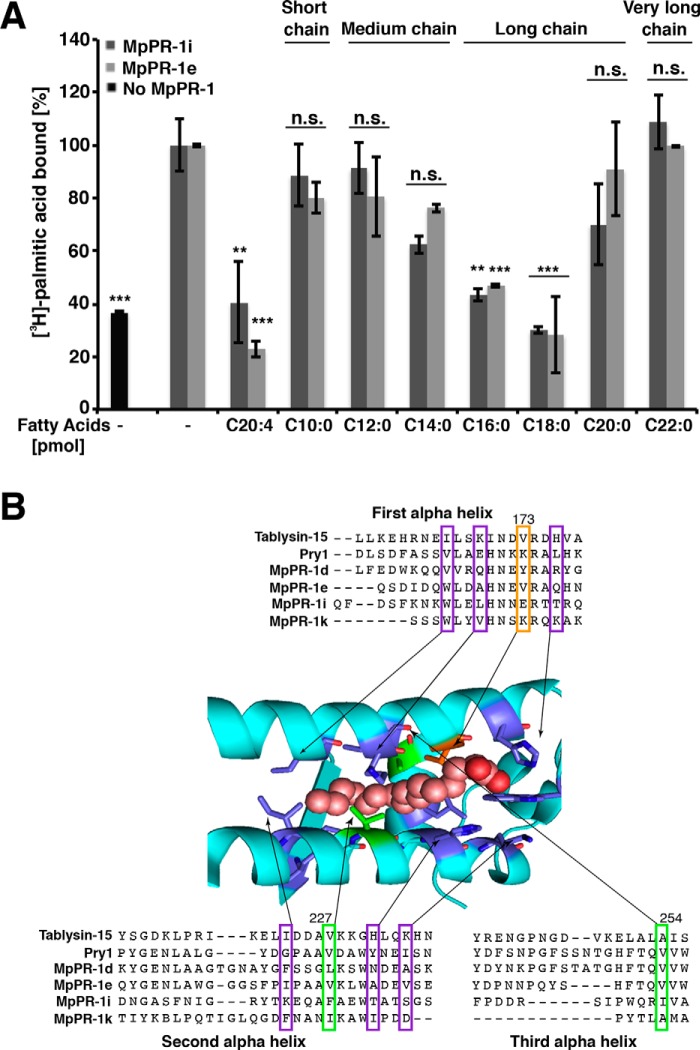
Moniliophthora perniciosa is the causative agent of witches' broom disease, which devastates cacao cultures in South America. This pathogenic fungus infects meristematic tissues and derives nutrients from the plant apoplast during an unusually long-lasting biotrophic stage. To survive, the fungus produces proteins to suppress the plant immune response. Proteins of the PR-1 (pathogenesis-related 1)/CAP superfamily have been implicated in fungal virulence and immune suppression. The genome of M. perniciosa encodes 11 homologues of plant PR-1 proteins, designated MpPR-1 proteins, but their precise mode of action is poorly understood. In this study, we expressed MpPR-1 proteins in a yeast model lacking endogenous CAP proteins. We show that some members of the MpPR-1 family bind and promote secretion of sterols, whereas others bind and promote secretion of fatty acids. Lipid binding by purified MpPR-1 occurs with micromolar affinity and is saturable in vitro Sterol binding by MpPR-1 requires the presence of a flexible loop region containing aromatic amino acids, the caveolin-binding motif. Remarkably, MpPR-1 family members that do not bind sterols can be converted to sterol binders by a single point mutation in the caveolin-binding motif. We discuss the possible implications of the lipid-binding activity of MpPR-1 family members with regard to the mode of action of these proteins during M. perniciosa infections.
Keywords: fatty acid; fatty acid binding protein; infection; lipid-protein interaction; sterol.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures

Similar articles
-
Crystal Structure of MpPR-1i, a SCP/TAPS protein from Moniliophthora perniciosa, the fungus that causes Witches' Broom Disease of Cacao.Sci Rep. 2017 Aug 10;7(1):7818. doi: 10.1038/s41598-017-07887-1. Sci Rep. 2017. PMID: 28798297 Free PMC article.
-
The fungal pathogen Moniliophthora perniciosa has genes similar to plant PR-1 that are highly expressed during its interaction with cacao.PLoS One. 2012;7(9):e45929. doi: 10.1371/journal.pone.0045929. Epub 2012 Sep 20. PLoS One. 2012. PMID: 23029323 Free PMC article.
-
The crystal structure of necrosis- and ethylene-inducing protein 2 from the causal agent of cacao's Witches' Broom disease reveals key elements for its activity.Biochemistry. 2011 Nov 15;50(45):9901-10. doi: 10.1021/bi201253b. Epub 2011 Oct 21. Biochemistry. 2011. PMID: 21999603
-
Moniliophthora perniciosa, the causal agent of witches' broom disease of cacao: what's new from this old foe?Mol Plant Pathol. 2008 Sep;9(5):577-88. doi: 10.1111/j.1364-3703.2008.00496.x. Mol Plant Pathol. 2008. PMID: 19018989 Free PMC article. Review.
-
Potential of Yeasts as Biocontrol Agents of the Phytopathogen Causing Cacao Witches' Broom Disease: Is Microbial Warfare a Solution?Front Microbiol. 2019 Jul 31;10:1766. doi: 10.3389/fmicb.2019.01766. eCollection 2019. Front Microbiol. 2019. PMID: 31417539 Free PMC article. Review.
Cited by
-
Adaptive evolution of Moniliophthora PR-1 proteins towards its pathogenic lifestyle.BMC Ecol Evol. 2021 May 14;21(1):84. doi: 10.1186/s12862-021-01818-5. BMC Ecol Evol. 2021. PMID: 33990179 Free PMC article.
-
State of the Art of the Molecular Biology of the Interaction between Cocoa and Witches' Broom Disease: A Systematic Review.Int J Mol Sci. 2023 Mar 16;24(6):5684. doi: 10.3390/ijms24065684. Int J Mol Sci. 2023. PMID: 36982760 Free PMC article.
-
Localization and functional characterization of the pathogenesis-related proteins Rbe1p and Rbt4p in Candida albicans.PLoS One. 2018 Aug 6;13(8):e0201932. doi: 10.1371/journal.pone.0201932. eCollection 2018. PLoS One. 2018. PMID: 30080909 Free PMC article.
-
Vertical and horizontal gene transfer shaped plant colonization and biomass degradation in the fungal genus Armillaria.Nat Microbiol. 2023 Sep;8(9):1668-1681. doi: 10.1038/s41564-023-01448-1. Epub 2023 Aug 7. Nat Microbiol. 2023. PMID: 37550506 Free PMC article.
-
Foliar Pine Pathogens From Different Kingdoms Share Defence-Eliciting Effector Proteins.Mol Plant Pathol. 2025 Mar;26(3):e70065. doi: 10.1111/mpp.70065. Mol Plant Pathol. 2025. PMID: 40025648 Free PMC article.
References
-
- Aime M. C., and Phillips-Mora W. (2005) The causal agents of witches' broom and frosty pod rot of cacao (chocolate, Theobroma cacao) form a new lineage of Marasmiaceae. Mycologia 97, 1012–1022 - PubMed
-
- Teixeira P. J., Thomazella D. P., Reis O., do Prado P. F., do Rio M. C., Fiorin G. L., José J., Costa G. G., Negri V. A., Mondego J. M., Mieczkowski P., and Pereira G. A. (2014) High-resolution transcript profiling of the atypical biotrophic interaction between Theobroma cacao and the fungal pathogen Moniliophthora perniciosa. Plant Cell 26, 4245–4269 - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
