

Unraveling the Self-Assembly of the Pseudomonas aeruginosa XcpQ Secretin Periplasmic Domain Provides New Molecular Insights into Type II Secretion System Secreton Architecture and Dynamics
- PMID: 29042493
- PMCID: PMC5646246
- DOI: 10.1128/mBio.01185-17
Unraveling the Self-Assembly of the Pseudomonas aeruginosa XcpQ Secretin Periplasmic Domain Provides New Molecular Insights into Type II Secretion System Secreton Architecture and Dynamics
Abstract
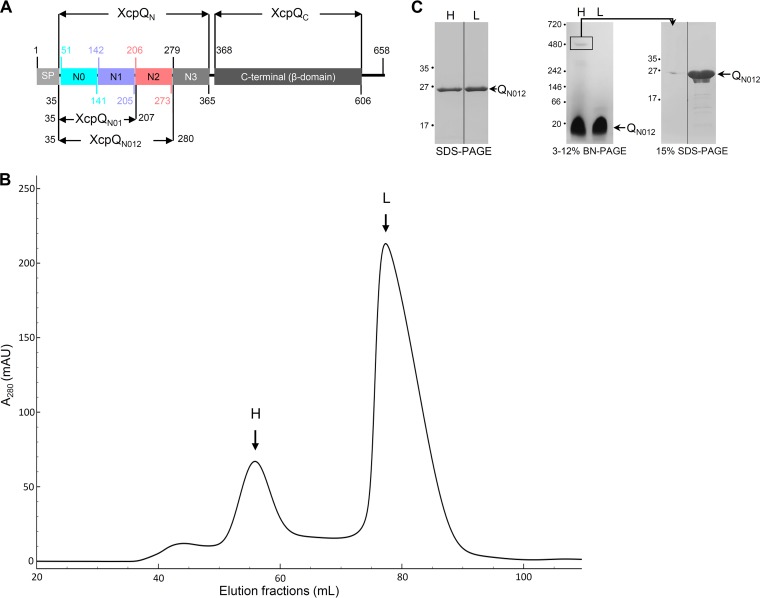
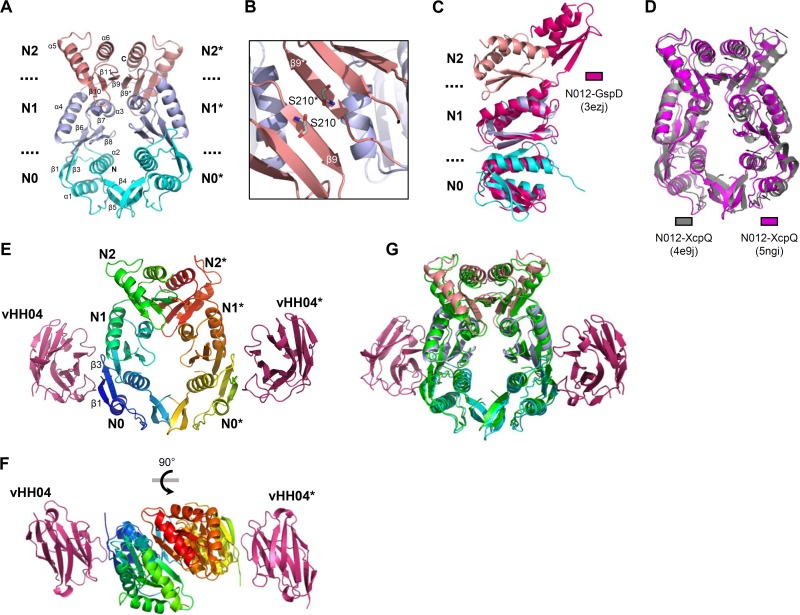
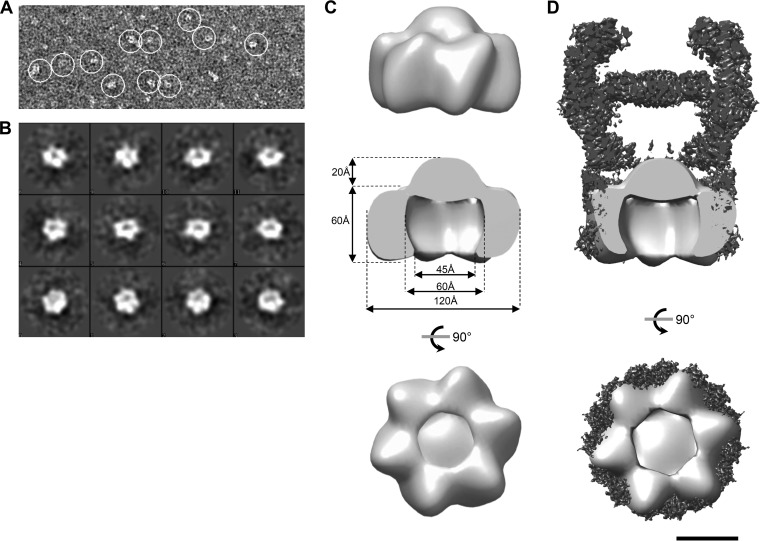
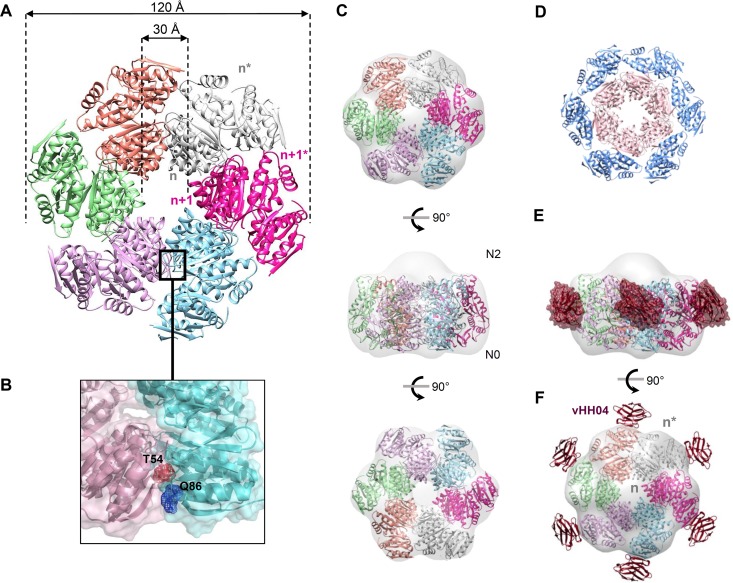
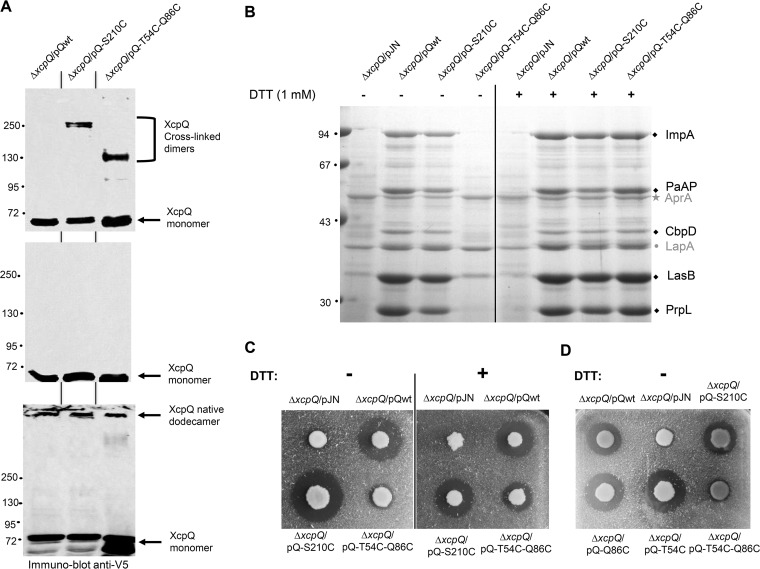
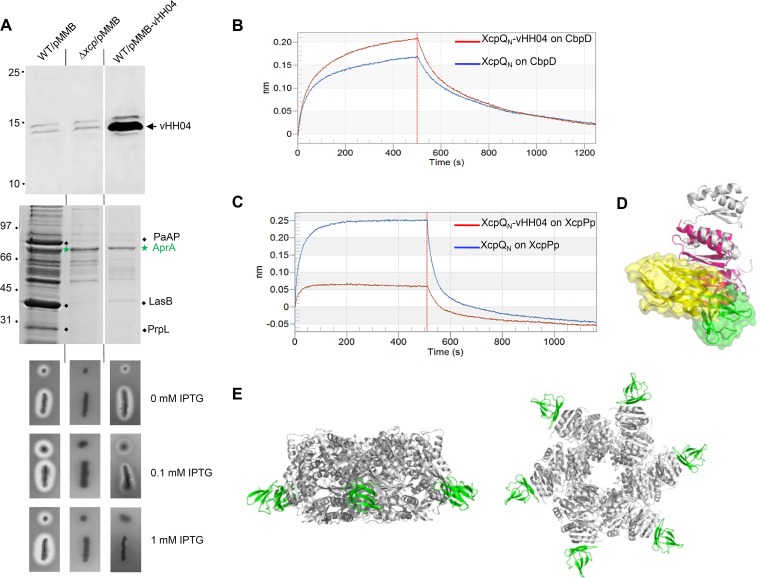
The type II secretion system (T2SS) releases large folded exoproteins across the envelope of many Gram-negative pathogens. This secretion process therefore requires specific gating, interacting, and dynamics properties mainly operated by a bipartite outer membrane channel called secretin. We have a good understanding of the structure-function relationship of the pore-forming C-terminal domain of secretins. In contrast, the high flexibility of their periplasmic N-terminal domain has been an obstacle in obtaining the detailed structural information required to uncover its molecular function. In Pseudomonas aeruginosa, the Xcp T2SS plays an important role in bacterial virulence by its capacity to deliver a large panel of toxins and degradative enzymes into the surrounding environment. Here, we revealed that the N-terminal domain of XcpQ secretin spontaneously self-assembled into a hexamer of dimers independently of its C-terminal domain. Furthermore, and by using multidisciplinary approaches, we elucidate the structural organization of the XcpQ N domain and demonstrate that secretin flexibility at interdimer interfaces is mandatory for its function.IMPORTANCE Bacterial secretins are large homooligomeric proteins constituting the outer membrane pore-forming element of several envelope-embedded nanomachines essential in bacterial survival and pathogenicity. They comprise a well-defined membrane-embedded C-terminal domain and a modular periplasmic N-terminal domain involved in substrate recruitment and connection with inner membrane components. We are studying the XcpQ secretin of the T2SS present in the pathogenic bacterium Pseudomonas aeruginosa Our data highlight the ability of the XcpQ N-terminal domain to spontaneously oligomerize into a hexamer of dimers. Further in vivo experiments revealed that this domain adopts different conformations essential for the T2SS secretion process. These findings provide new insights into the functional understanding of bacterial T2SS secretins.
Keywords: Pseudomonas aeruginosa; dynamics; protein structure-function; secretin; stoichiometry; type II secretion system.
Copyright © 2017 Douzi et al.
Figures
Similar articles
-
Structure and assembly of pilotin-dependent and -independent secretins of the type II secretion system.PLoS Pathog. 2019 May 13;15(5):e1007731. doi: 10.1371/journal.ppat.1007731. eCollection 2019 May. PLoS Pathog. 2019. PMID: 31083688 Free PMC article.
-
Structural Basis of Type 2 Secretion System Engagement between the Inner and Outer Bacterial Membranes.mBio. 2017 Oct 17;8(5):e01344-17. doi: 10.1128/mBio.01344-17. mBio. 2017. PMID: 29042496 Free PMC article.
-
New insights into the assembly of bacterial secretins: structural studies of the periplasmic domain of XcpQ from Pseudomonas aeruginosa.J Biol Chem. 2013 Jan 11;288(2):1214-25. doi: 10.1074/jbc.M112.432096. Epub 2012 Nov 27. J Biol Chem. 2013. PMID: 23188826 Free PMC article.
-
Architecture, Function, and Substrates of the Type II Secretion System.EcoSal Plus. 2019 Feb;8(2):10.1128/ecosalplus.ESP-0034-2018. doi: 10.1128/ecosalplus.ESP-0034-2018. EcoSal Plus. 2019. PMID: 30767847 Free PMC article. Review.
-
The trans-envelope architecture and function of the type 2 secretion system: new insights raising new questions.Mol Microbiol. 2017 Jul;105(2):211-226. doi: 10.1111/mmi.13704. Epub 2017 May 18. Mol Microbiol. 2017. PMID: 28486768 Review.
Cited by
-
Structure and assembly of pilotin-dependent and -independent secretins of the type II secretion system.PLoS Pathog. 2019 May 13;15(5):e1007731. doi: 10.1371/journal.ppat.1007731. eCollection 2019 May. PLoS Pathog. 2019. PMID: 31083688 Free PMC article.
-
Direct interactions between the secreted effector and the T2SS components GspL and GspM reveal a new effector-sensing step during type 2 secretion.J Biol Chem. 2018 Dec 14;293(50):19441-19450. doi: 10.1074/jbc.RA117.001127. Epub 2018 Oct 18. J Biol Chem. 2018. PMID: 30337370 Free PMC article.
-
Insights into dynamics and gating properties of T2SS secretins.Sci Adv. 2023 Oct 6;9(40):eadg6996. doi: 10.1126/sciadv.adg6996. Epub 2023 Oct 4. Sci Adv. 2023. PMID: 37792935 Free PMC article.
-
Differential Modulation of Quorum Sensing Signaling through QslA in Pseudomonas aeruginosa Strains PAO1 and PA14.J Bacteriol. 2019 Oct 4;201(21):e00362-19. doi: 10.1128/JB.00362-19. Print 2019 Nov 1. J Bacteriol. 2019. PMID: 31405911 Free PMC article.
-
Bacterial secretins: Mechanisms of assembly and membrane targeting.Protein Sci. 2020 Apr;29(4):893-904. doi: 10.1002/pro.3835. Epub 2020 Feb 19. Protein Sci. 2020. PMID: 32020694 Free PMC article. Review.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
