

Developmental stage-dependent regulation of spine formation by calcium-calmodulin-dependent protein kinase IIα and Rap1
- PMID: 29042611
- PMCID: PMC5645322
- DOI: 10.1038/s41598-017-13728-y
Developmental stage-dependent regulation of spine formation by calcium-calmodulin-dependent protein kinase IIα and Rap1
Abstract
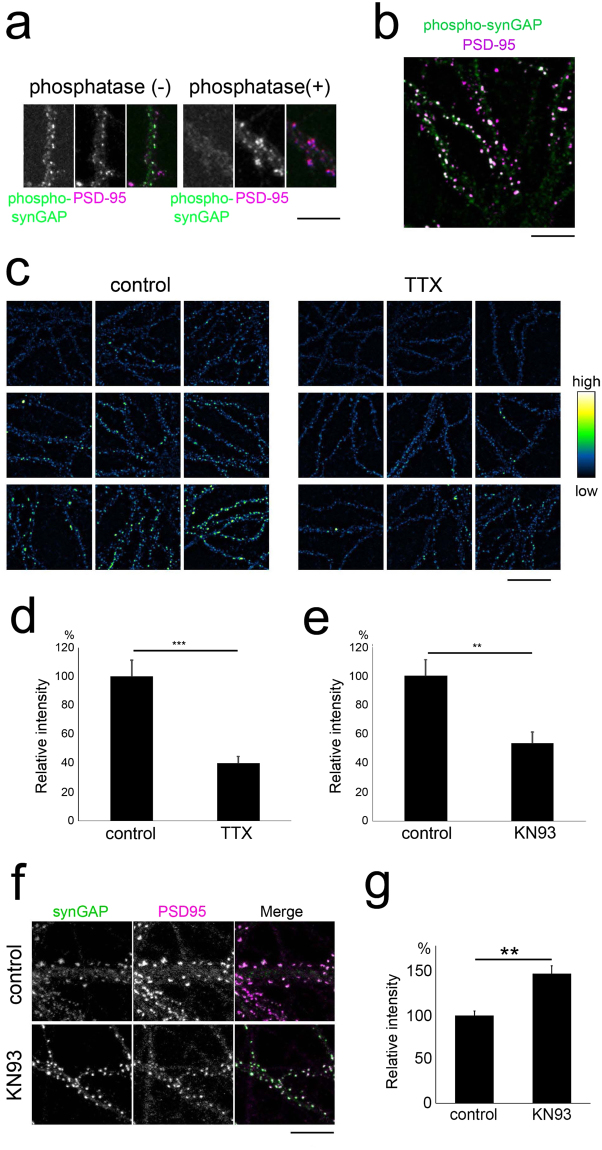
The roles of calcium-calmodulin-dependent protein kinase II-alpha (CaMKIIα) in the expression of long-term synaptic plasticity in the adult brain have been extensively studied. However, how increased CaMKIIα activity controls the maturation of neuronal circuits remains incompletely understood. Herein, we show that pyramidal neurons without CaMKIIα activity upregulate the rate of spine addition, resulting in elevated spine density. Genetic elimination of CaMKIIα activity specifically eliminated the observed maturation-dependent suppression of spine formation. Enhanced spine formation was associated with the stabilization of actin in the spine and could be reversed by increasing the activity of the small GTPase Rap1. CaMKIIα activity was critical in the phosphorylation of synaptic Ras GTPase-activating protein (synGAP), the dispersion of synGAP from postsynaptic sites, and the activation of postsynaptic Rap1. CaMKIIα is already known to be essential in learning and memory, but our findings suggest that CaMKIIα plays an important activity-dependent role in restricting spine density during postnatal development.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures







Similar articles
-
Phosphorylation of synaptic GTPase-activating protein (synGAP) by Ca2+/calmodulin-dependent protein kinase II (CaMKII) and cyclin-dependent kinase 5 (CDK5) alters the ratio of its GAP activity toward Ras and Rap GTPases.J Biol Chem. 2015 Feb 20;290(8):4908-4927. doi: 10.1074/jbc.M114.614420. Epub 2014 Dec 22. J Biol Chem. 2015. PMID: 25533468 Free PMC article.
-
Kinase-dead knock-in mouse reveals an essential role of kinase activity of Ca2+/calmodulin-dependent protein kinase IIalpha in dendritic spine enlargement, long-term potentiation, and learning.J Neurosci. 2009 Jun 10;29(23):7607-18. doi: 10.1523/JNEUROSCI.0707-09.2009. J Neurosci. 2009. PMID: 19515929 Free PMC article.
-
Activity-dependent modulation of the interaction between CaMKIIα and Abi1 and its involvement in spine maturation.J Neurosci. 2012 Sep 19;32(38):13177-88. doi: 10.1523/JNEUROSCI.2257-12.2012. J Neurosci. 2012. PMID: 22993434 Free PMC article.
-
Stimulation-induced changes in diffusion and structure of calmodulin and calmodulin-dependent protein kinase II proteins in neurons.Neurosci Res. 2018 Nov;136:13-32. doi: 10.1016/j.neures.2018.01.003. Epub 2018 Feb 1. Neurosci Res. 2018. PMID: 29395358 Review.
-
Structural modulation of dendritic spines during synaptic plasticity.Neuroscientist. 2012 Aug;18(4):326-41. doi: 10.1177/1073858411407206. Epub 2011 Jun 13. Neuroscientist. 2012. PMID: 21670426 Review.
Cited by
-
The ISR downstream target ATF4 represses long-term memory in a cell type-specific manner.Proc Natl Acad Sci U S A. 2024 Jul 30;121(31):e2407472121. doi: 10.1073/pnas.2407472121. Epub 2024 Jul 24. Proc Natl Acad Sci U S A. 2024. PMID: 39047038 Free PMC article.
-
Physics Clues on the Mind Substrate and Attributes.Front Comput Neurosci. 2022 Apr 8;16:836532. doi: 10.3389/fncom.2022.836532. eCollection 2022. Front Comput Neurosci. 2022. PMID: 35465268 Free PMC article.
-
Activity-regulated E3 ubiquitin ligase TRIM47 modulates excitatory synapse development.Front Mol Neurosci. 2022 Sep 21;15:943980. doi: 10.3389/fnmol.2022.943980. eCollection 2022. Front Mol Neurosci. 2022. PMID: 36211980 Free PMC article.
-
The dopamine analogue CA140 alleviates AD pathology, neuroinflammation, and rescues synaptic/cognitive functions by modulating DRD1 signaling or directly binding to Abeta.J Neuroinflammation. 2024 Aug 11;21(1):200. doi: 10.1186/s12974-024-03180-x. J Neuroinflammation. 2024. PMID: 39129007 Free PMC article.
-
Stress-mediated dysregulation of the Rap1 small GTPase impairs hippocampal structure and function.iScience. 2023 Aug 7;26(9):107566. doi: 10.1016/j.isci.2023.107566. eCollection 2023 Sep 15. iScience. 2023. PMID: 37664580 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
