

Enterococci Mediate the Oviposition Preference of Drosophila melanogaster through Sucrose Catabolism
- PMID: 29044155
- PMCID: PMC5647369
- DOI: 10.1038/s41598-017-13705-5
Enterococci Mediate the Oviposition Preference of Drosophila melanogaster through Sucrose Catabolism
Abstract
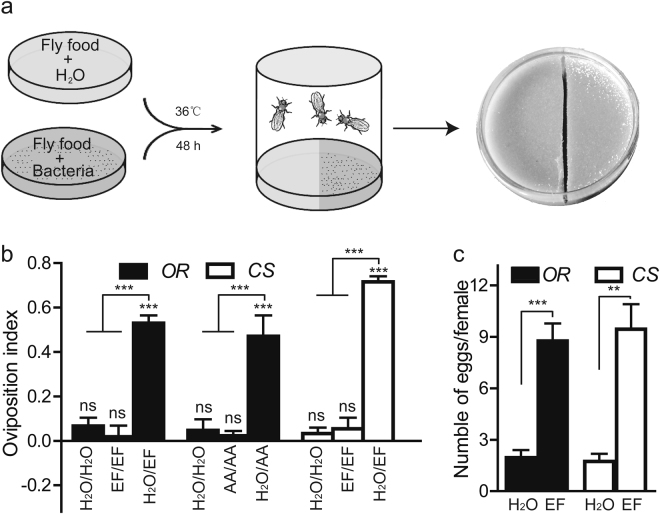
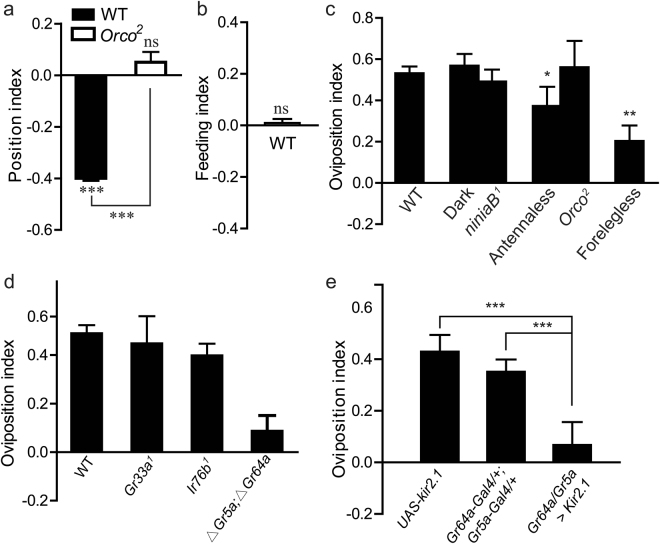
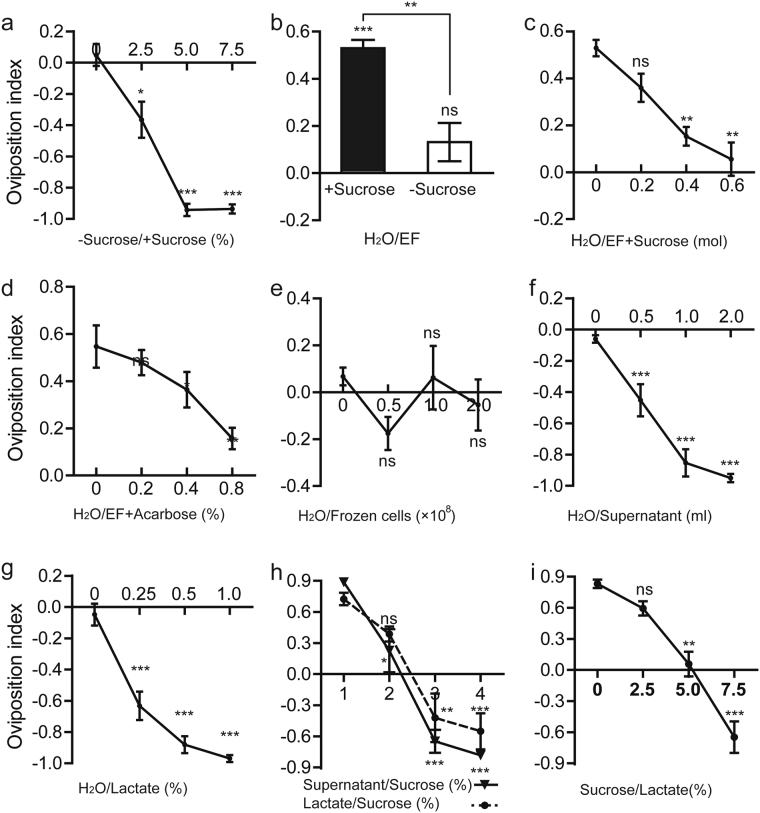
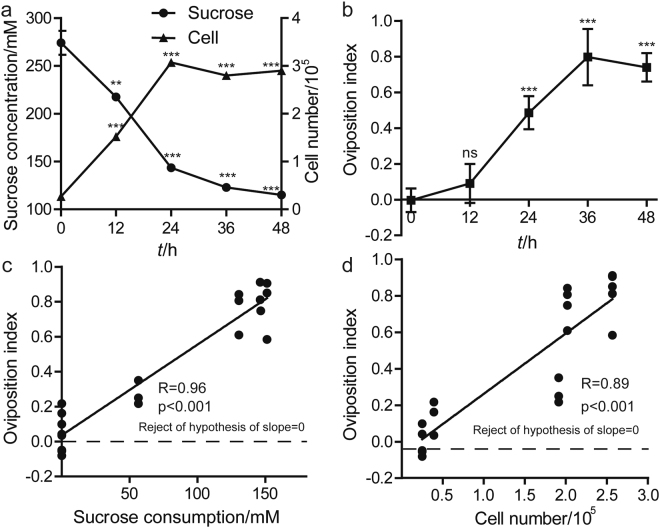
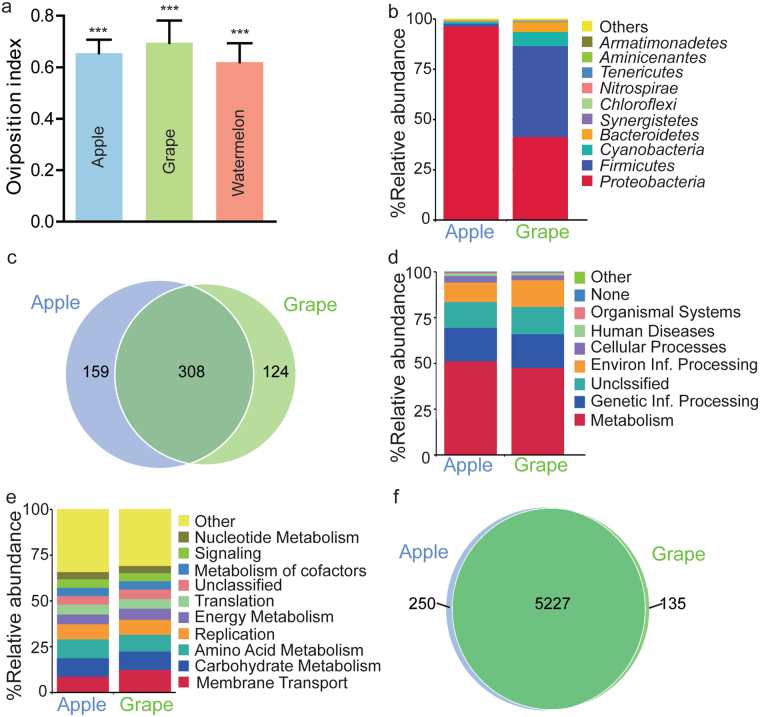
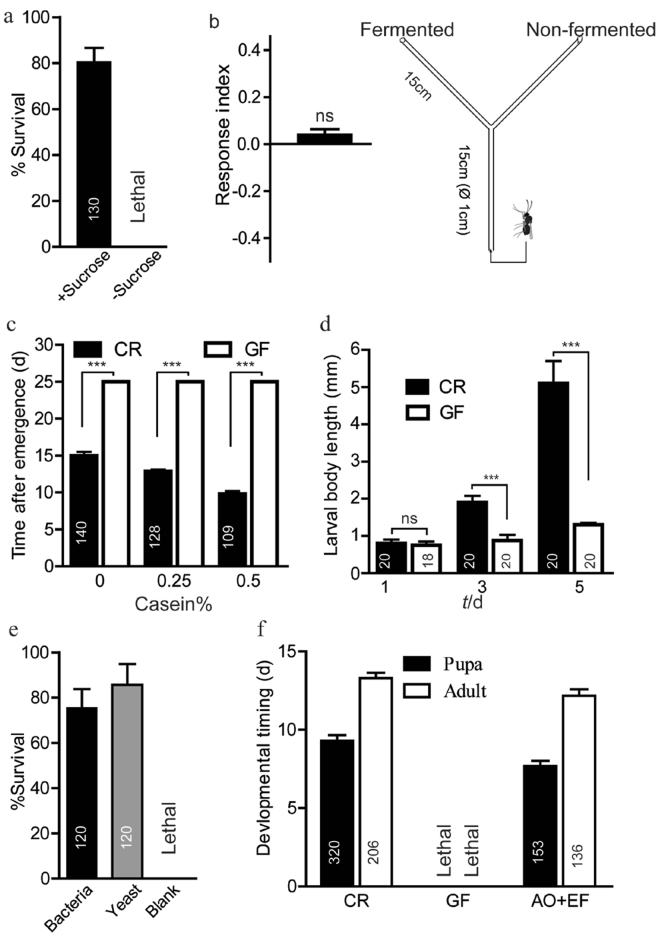
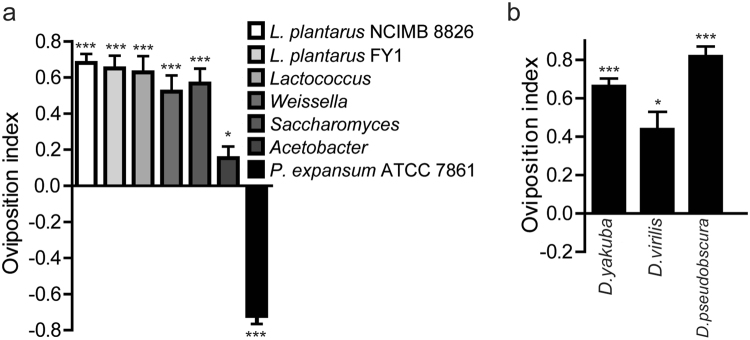
Sucrose, one of the main products of photosynthesis in plants, functions as a universal biomarker for nutritional content and maturity of different fruits across diverse ecological niches. Drosophila melanogaster congregates to lay eggs in rotting fruits, yet the factors that influence these decisions remains uncovered. Here, we report that lactic acid bacteria Enterococci are critical modulators to attract Drosophila to lay eggs on decaying food. Drosophila-associated Enterococci predominantly catabolize sucrose for growing their population in fly food, and thus generate a unique ecological niche with depleted sucrose, but enriched bacteria. Female flies navigate these favorable oviposition sites by probing the sucrose cue with their gustatory sensory neurons. Acquirement of indigenous microbiota facilitated the development and systemic growth of Drosophila, thereby benefiting the survival and fitness of their offspring. Thus, our finding highlights the pivotal roles of commensal bacteria in influencing host behavior, opening the door to a better understanding of the ecological relationships between the microbial and metazoan worlds.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Ezenwa VO, Gerardo NM, Inouye DW, Medina M, Xavier JB. Microbiology. Animal behavior and the microbiome. Science. 2012;338:198–199. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
