

Regulation of valve interstitial cell homeostasis by mechanical deformation: implications for heart valve disease and surgical repair
- PMID: 29046338
- PMCID: PMC5665836
- DOI: 10.1098/rsif.2017.0580
Regulation of valve interstitial cell homeostasis by mechanical deformation: implications for heart valve disease and surgical repair
Abstract
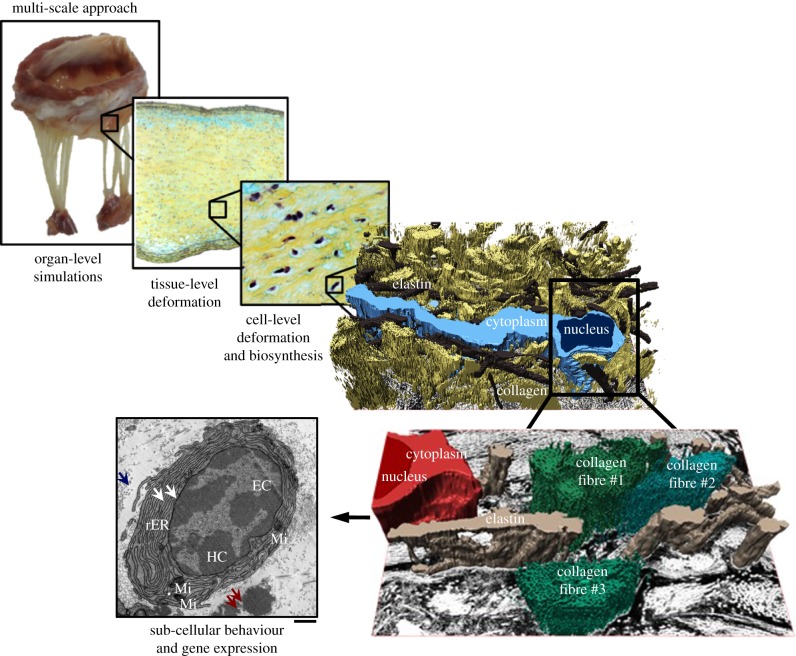
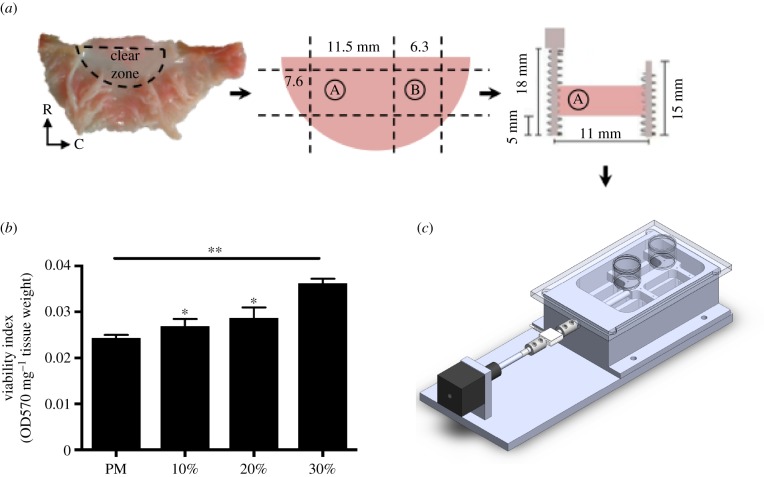
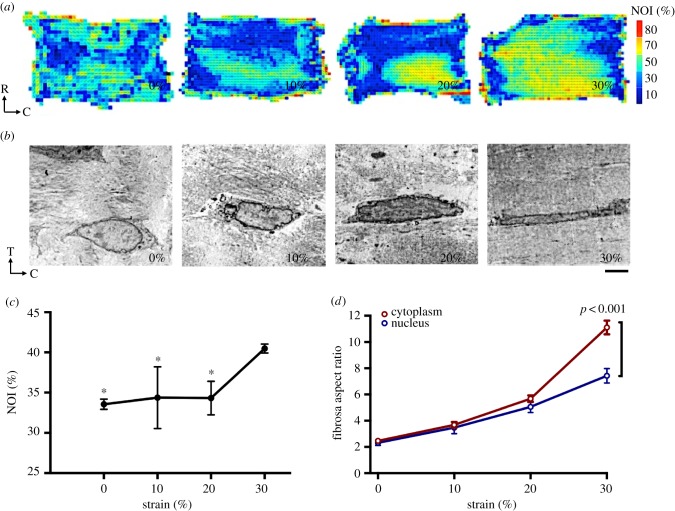
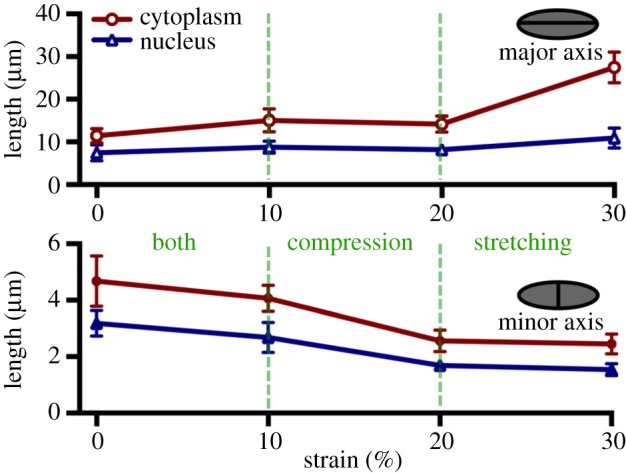
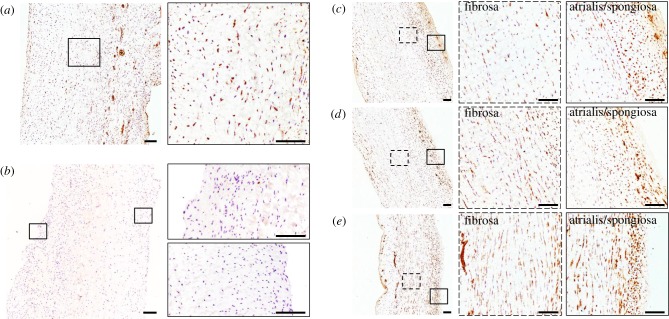
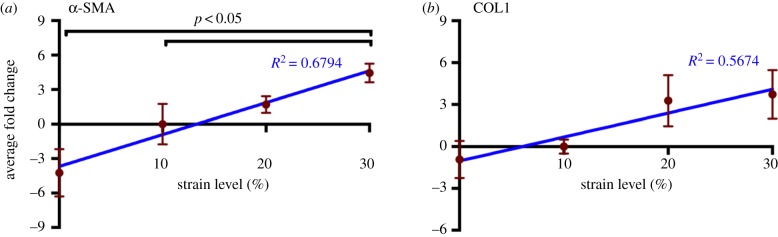
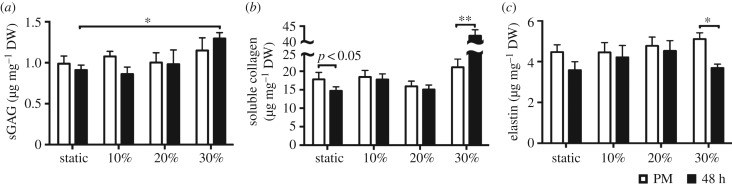
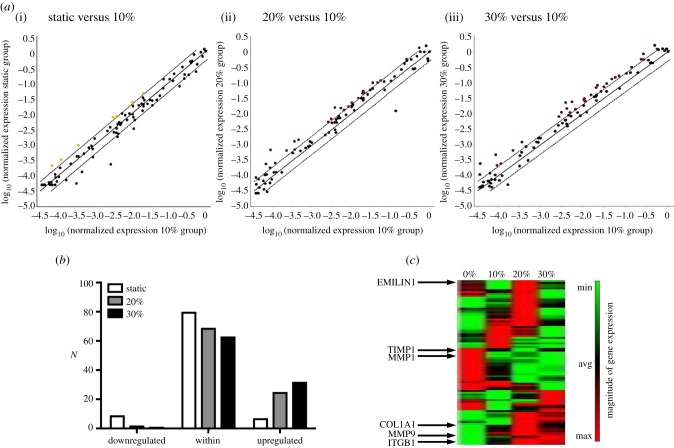
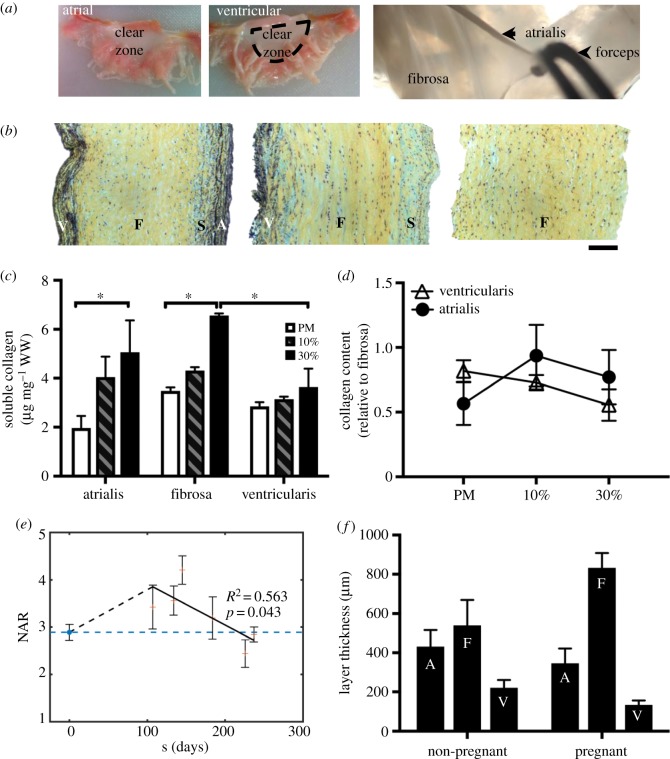
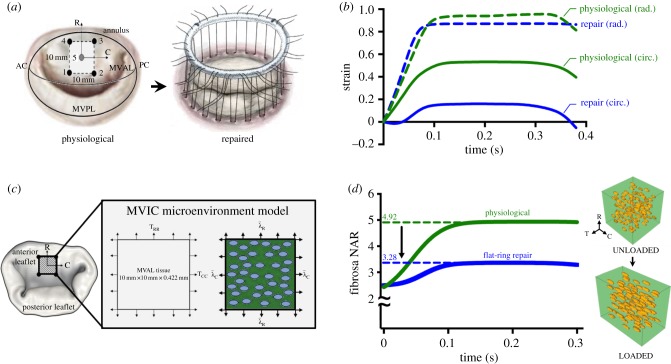
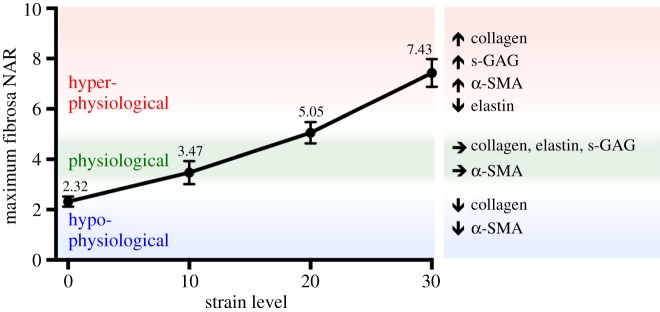
Mechanical stress is one of the major aetiological factors underlying soft-tissue remodelling, especially for the mitral valve (MV). It has been hypothesized that altered MV tissue stress states lead to deviations from cellular homeostasis, resulting in subsequent cellular activation and extracellular matrix (ECM) remodelling. However, a quantitative link between alterations in the organ-level in vivo state and in vitro-based mechanobiology studies has yet to be made. We thus developed an integrated experimental-computational approach to elucidate MV tissue and interstitial cell responses to varying tissue strain levels. Comprehensive results at different length scales revealed that normal responses are observed only within a defined range of tissue deformations, whereas deformations outside of this range lead to hypo- and hyper-synthetic responses, evidenced by changes in α-smooth muscle actin, type I collagen, and other ECM and cell adhesion molecule regulation. We identified MV interstitial cell deformation as a key player in leaflet tissue homeostatic regulation and, as such, used it as the metric that makes the critical link between in vitro responses to simulated equivalent in vivo behaviour. Results indicated that cell responses have a delimited range of in vivo deformations that maintain a homeostatic response, suggesting that deviations from this range may lead to deleterious tissue remodelling and failure.
Keywords: collagen; myofibroblast; valve interstitial cells.
© 2017 The Author(s).
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Ogden R. 2003. Nonlinear elasticity, anisotropy, material stability, and residual stresses in soft tissue. In Biomechanics of soft tissue in cardiovascular system (ed. Ogden RW.), pp. 65–108. New York, NY: Springer.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
