

Myeloid Dendritic Cells Repress Human Cytomegalovirus Gene Expression and Spread by Releasing Interferon-Unrelated Soluble Antiviral Factors
- PMID: 29046460
- PMCID: PMC5730771
- DOI: 10.1128/JVI.01138-17
Myeloid Dendritic Cells Repress Human Cytomegalovirus Gene Expression and Spread by Releasing Interferon-Unrelated Soluble Antiviral Factors
Abstract
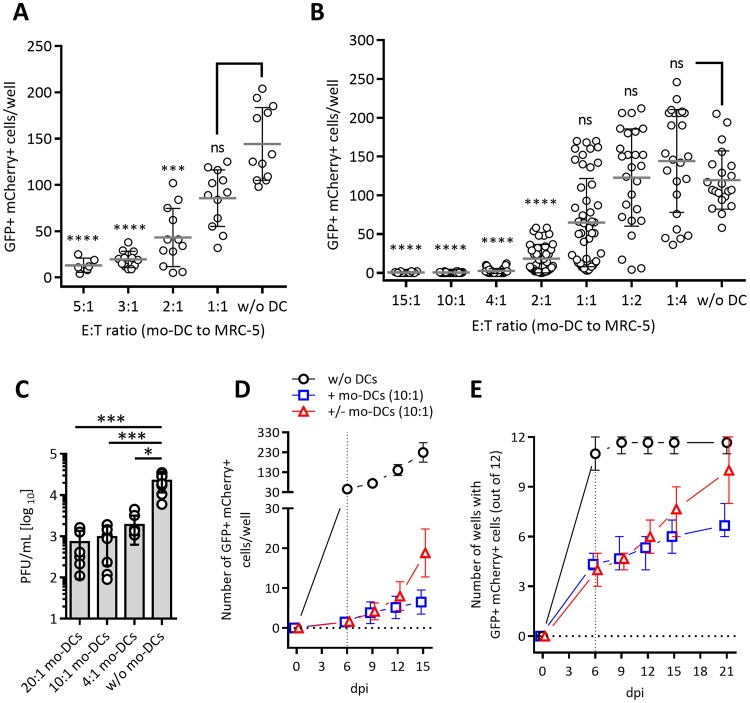
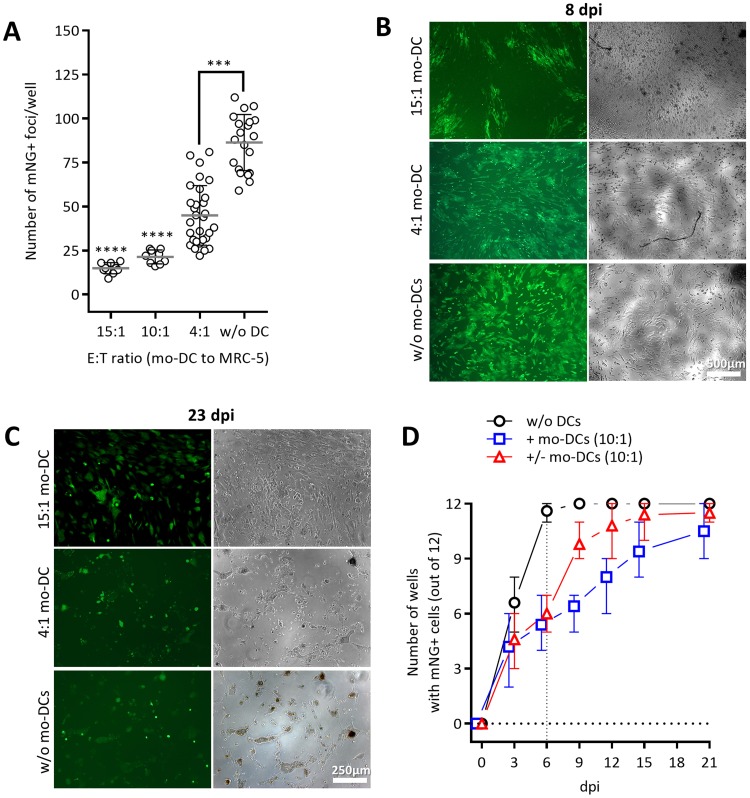
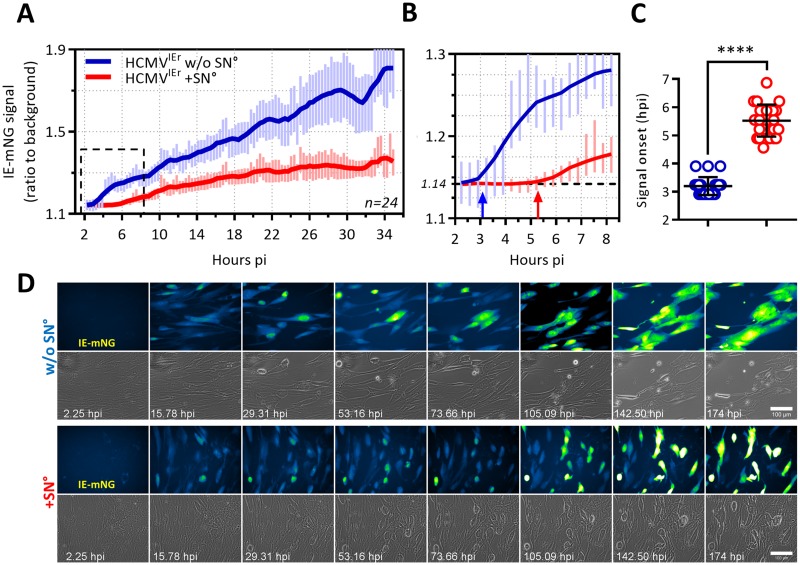
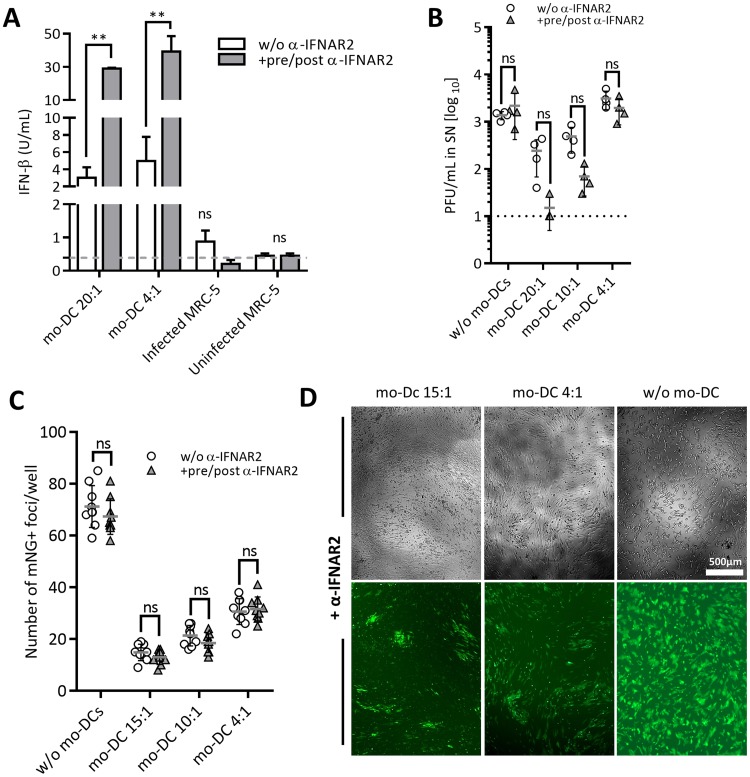
Cytomegalovirus (CMV) is a betaherpesvirus that latently infects most adult humans worldwide and is a major cause of morbidity and mortality in immunocompromised hosts. Latent human CMV (HCMV) is believed to reside in precursors of myeloid-lineage leukocytes and monocytes, which give rise to macrophages and dendritic cells (DC). We report here that human monocyte-derived DC (mo-DC) suppress HCMV infection in coculture with infected fibroblast target cells in a manner dependent on the effector-to-target ratio. Intriguingly, optimal activation of mo-DC was achieved under coculture conditions and not by direct infection with HCMV, implying that mo-DC may recognize unique molecular patterns on, or within, infected fibroblasts. We show that HCMV is controlled by secreted factors that act by priming defenses in target cells rather than by direct viral neutralization, but we excluded a role for interferons (IFNs) in this control. The expression of lytic viral genes in infected cells and the progression of infection were significantly slowed, but this effect was reversible, indicating that the control of infection depended on the transient induction of antiviral effector molecules in target cells. Using immediate early or late-phase reporter HCMVs, we show that soluble factors secreted in the cocultures suppress HCMV replication at both stages of the infection and that their antiviral effects are robust and comparable in numerous batches of mo-DC as well as in primary fibroblasts and stromal cells.IMPORTANCE Human cytomegalovirus is a widespread opportunistic pathogen that can cause severe disease and complications in vulnerable individuals. This includes newborn children, HIV AIDS patients, and transplant recipients. Although the majority of healthy humans carry this virus throughout their lives without symptoms, it is not exactly clear which tissues in the body are the main reservoirs of latent virus infection or how the delicate balance between the virus and the immune system is maintained over an individual's lifetime. Here, for the first time, we provide evidence for a novel mechanism of direct virus control by a subset of human innate immune cells called dendritic cells, which are regarded as a major site of virus latency and reactivation. Our findings may have important implications in HCMV disease prevention as well as in development of novel therapeutic approaches.
Keywords: antiviral agents; cytomegalovirus; dendritic cells; fluorescent image analysis; video microscopy.
Copyright © 2017 American Society for Microbiology.
Figures
Similar articles
-
Type I Interferon Released by Myeloid Dendritic Cells Reversibly Impairs Cytomegalovirus Replication by Inhibiting Immediate Early Gene Expression.J Virol. 2015 Oct;89(19):9886-95. doi: 10.1128/JVI.01459-15. Epub 2015 Jul 22. J Virol. 2015. PMID: 26202227 Free PMC article.
-
Human cytomegalovirus modulates monocyte-mediated innate immune responses during short-term experimental latency in vitro.J Virol. 2014 Aug;88(16):9391-405. doi: 10.1128/JVI.00934-14. Epub 2014 Jun 11. J Virol. 2014. PMID: 24920803 Free PMC article.
-
Latency-Associated Expression of Human Cytomegalovirus US28 Attenuates Cell Signaling Pathways To Maintain Latent Infection.mBio. 2017 Dec 5;8(6):e01754-17. doi: 10.1128/mBio.01754-17. mBio. 2017. PMID: 29208743 Free PMC article.
-
Human cytomegalovirus: Latency and reactivation in the myeloid lineage.J Clin Virol. 2008 Mar;41(3):180-5. doi: 10.1016/j.jcv.2007.11.014. J Clin Virol. 2008. PMID: 18164651 Review.
-
Aspects of human cytomegalovirus latency and reactivation.Curr Top Microbiol Immunol. 2008;325:297-313. doi: 10.1007/978-3-540-77349-8_17. Curr Top Microbiol Immunol. 2008. PMID: 18637513 Review.
Cited by
-
Organoid modeling of Zika and herpes simplex virus 1 infections reveals virus-specific responses leading to microcephaly.Cell Stem Cell. 2021 Aug 5;28(8):1362-1379.e7. doi: 10.1016/j.stem.2021.03.004. Epub 2021 Apr 9. Cell Stem Cell. 2021. PMID: 33838105 Free PMC article.
-
A Novel Triple-Fluorescent HCMV Strain Reveals Gene Expression Dynamics and Anti-Herpesviral Drug Mechanisms.Front Cell Infect Microbiol. 2021 Jan 8;10:536150. doi: 10.3389/fcimb.2020.536150. eCollection 2020. Front Cell Infect Microbiol. 2021. PMID: 33489928 Free PMC article.
-
Cytomegalovirus inhibition of extrinsic apoptosis determines fitness and resistance to cytotoxic CD8 T cells.Proc Natl Acad Sci U S A. 2020 Jun 9;117(23):12961-12968. doi: 10.1073/pnas.1914667117. Epub 2020 May 22. Proc Natl Acad Sci U S A. 2020. PMID: 32444487 Free PMC article.
-
Human cytomegalovirus UL23 inhibits immune cell migration and blocks antiviral immune cell responses by reducing the expression of chemokines CCL2 and CCL5.Virulence. 2025 Dec;16(1):2500493. doi: 10.1080/21505594.2025.2500493. Epub 2025 Jun 16. Virulence. 2025. PMID: 40521898 Free PMC article.
-
Exogenous Interleukin-37 Alleviates Hepatitis with Reduced Dendritic Cells and Induced Regulatory T Cells in Acute Murine Cytomegalovirus Infection.J Immunol Res. 2023 May 12;2023:1462048. doi: 10.1155/2023/1462048. eCollection 2023. J Immunol Res. 2023. PMID: 37215069 Free PMC article.
References
-
- Boppana SB, Fowler KB. 2007. HCMV: persistence in the population: epidemiology and transmission, p 795–813. In Arvin A, Campadelli-Fiume C, Mocarski E, Moore PS, Roizamn B, Whitley R, Yamanishi K (ed), Human herpesviruses: biology, therapy, and immunoprophylaxis. Cambridge University Press, Cambridge, UK. - PubMed
-
- Söderberg-Nauclér C, Nelson JA. 1999. Human cytomegalovirus latency and reactivation–a delicate balance between the virus and its host's immune system. Intervirology 42:314–421. - PubMed
-
- Sylwester AW, Mitchell BL, Edgar JB, Taormina C, Pelte C, Ruchti F, Sleath PR, Grabstein KH, Hosken NA, Kern F, Nelson JA, Picker LJ. 2005. Broadly targeted human cytomegalovirus-specific CD4+ and CD8+ T cells dominate the memory compartments of exposed subjects. J Exp Med 202:673–685. doi:10.1084/jem.20050882. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
