

Interpreting T-Cell Cross-reactivity through Structure: Implications for TCR-Based Cancer Immunotherapy
- PMID: 29046675
- PMCID: PMC5632759
- DOI: 10.3389/fimmu.2017.01210
Interpreting T-Cell Cross-reactivity through Structure: Implications for TCR-Based Cancer Immunotherapy
Abstract
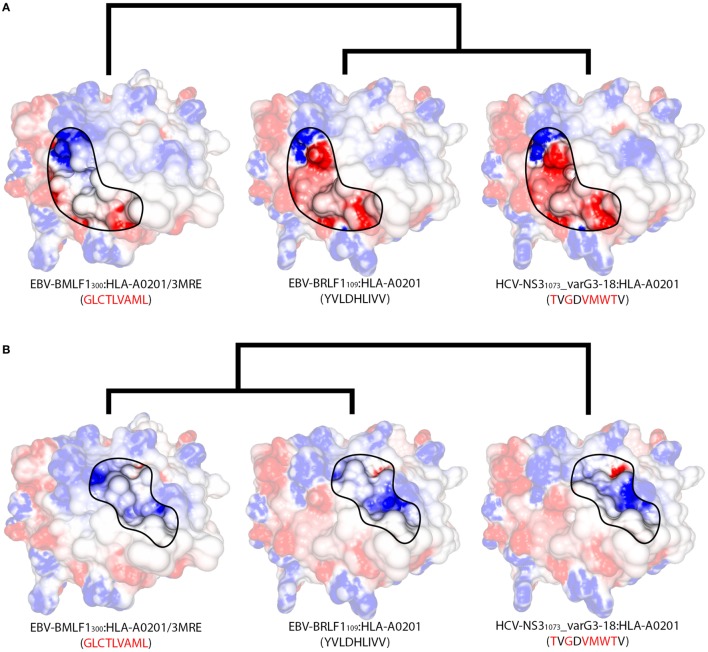
Immunotherapy has become one of the most promising avenues for cancer treatment, making use of the patient's own immune system to eliminate cancer cells. Clinical trials with T-cell-based immunotherapies have shown dramatic tumor regressions, being effective in multiple cancer types and for many different patients. Unfortunately, this progress was tempered by reports of serious (even fatal) side effects. Such therapies rely on the use of cytotoxic T-cell lymphocytes, an essential part of the adaptive immune system. Cytotoxic T-cells are regularly involved in surveillance and are capable of both eliminating diseased cells and generating protective immunological memory. The specificity of a given T-cell is determined through the structural interaction between the T-cell receptor (TCR) and a peptide-loaded major histocompatibility complex (MHC); i.e., an intracellular peptide-ligand displayed at the cell surface by an MHC molecule. However, a given TCR can recognize different peptide-MHC (pMHC) complexes, which can sometimes trigger an unwanted response that is referred to as T-cell cross-reactivity. This has become a major safety issue in TCR-based immunotherapies, following reports of melanoma-specific T-cells causing cytotoxic damage to healthy tissues (e.g., heart and nervous system). T-cell cross-reactivity has been extensively studied in the context of viral immunology and tissue transplantation. Growing evidence suggests that it is largely driven by structural similarities of seemingly unrelated pMHC complexes. Here, we review recent reports about the existence of pMHC "hot-spots" for cross-reactivity and propose the existence of a TCR interaction profile (i.e., a refinement of a more general TCR footprint in which some amino acid residues are more important than others in triggering T-cell cross-reactivity). We also make use of available structural data and pMHC models to interpret previously reported cross-reactivity patterns among virus-derived peptides. Our study provides further evidence that structural analyses of pMHC complexes can be used to assess the intrinsic likelihood of cross-reactivity among peptide-targets. Furthermore, we hypothesize that some apparent inconsistencies in reported cross-reactivities, such as a preferential directionality, might also be driven by particular structural features of the targeted pMHC complex. Finally, we explain why TCR-based immunotherapy provides a special context in which meaningful T-cell cross-reactivity predictions can be made.
Keywords: T-cell cross-reactivity; TCR-interacting surface; TCR/pMHC; cancer immunotherapy; cross-reactivity hot-spots; hierarchical clustering; peptide–MHC complex.
Figures
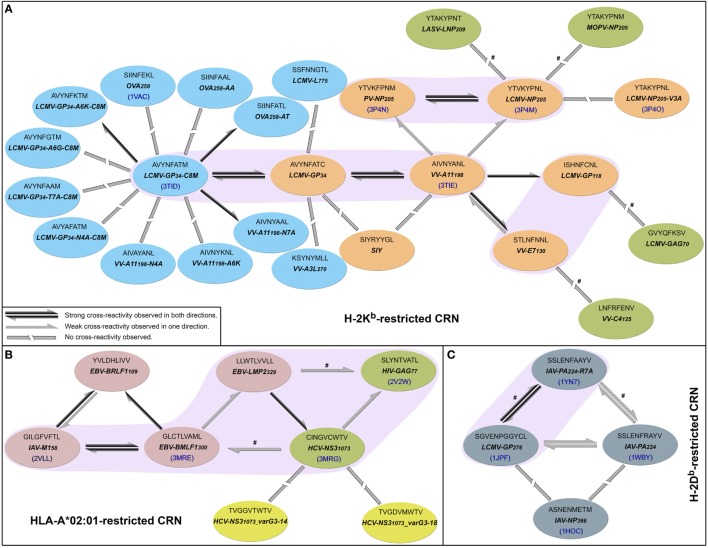
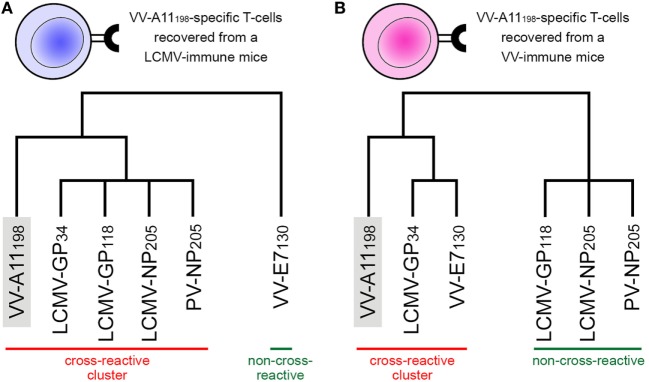
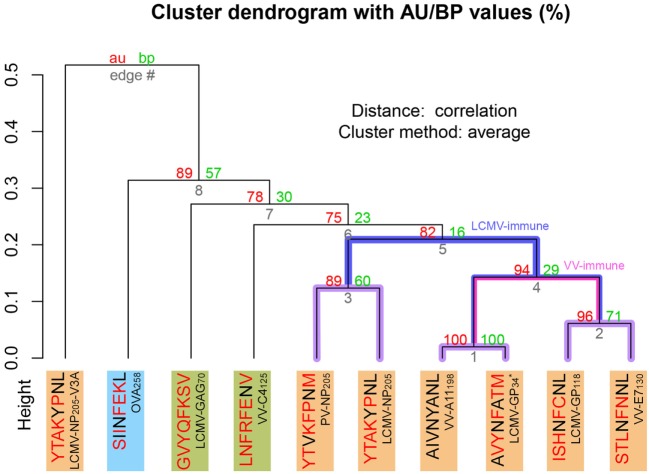
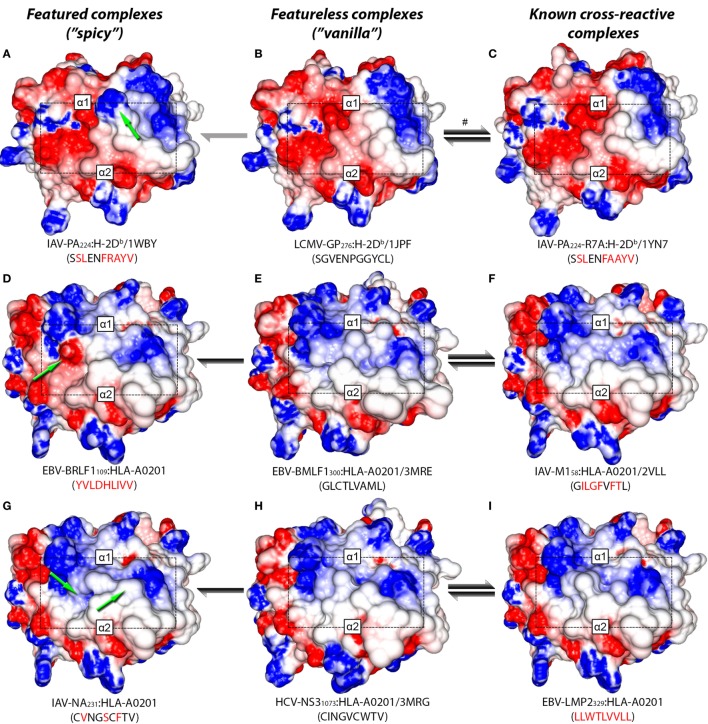
Similar articles
-
MPID-T: database for sequence-structure-function information on T-cell receptor/peptide/MHC interactions.Appl Bioinformatics. 2006;5(2):111-4. doi: 10.2165/00822942-200605020-00005. Appl Bioinformatics. 2006. PMID: 16722775
-
Improved structural method for T-cell cross-reactivity prediction.Mol Immunol. 2015 Oct;67(2 Pt B):303-10. doi: 10.1016/j.molimm.2015.06.017. Epub 2015 Jul 2. Mol Immunol. 2015. PMID: 26141239
-
A structural-based machine learning method to classify binding affinities between TCR and peptide-MHC complexes.Mol Immunol. 2021 Nov;139:76-86. doi: 10.1016/j.molimm.2021.07.020. Epub 2021 Aug 26. Mol Immunol. 2021. PMID: 34455212 Free PMC article.
-
Modulation of T cell function by TCR/pMHC binding kinetics.Immunobiology. 2006;211(1-2):47-64. doi: 10.1016/j.imbio.2005.09.003. Epub 2006 Jan 4. Immunobiology. 2006. PMID: 16446170 Review.
-
Structural and thermodynamic correlates of T cell signaling.Annu Rev Biophys Biomol Struct. 2002;31:121-49. doi: 10.1146/annurev.biophys.31.082901.134423. Epub 2001 Oct 25. Annu Rev Biophys Biomol Struct. 2002. PMID: 11988465 Review.
Cited by
-
Sequence-structure-function relationships in class I MHC: A local frustration perspective.PLoS One. 2020 May 18;15(5):e0232849. doi: 10.1371/journal.pone.0232849. eCollection 2020. PLoS One. 2020. PMID: 32421728 Free PMC article.
-
Mechanistic diversity in MHC class I antigen recognition.Biochem J. 2021 Dec 22;478(24):4187-4202. doi: 10.1042/BCJ20200910. Biochem J. 2021. PMID: 34940832 Free PMC article. Review.
-
MatchTope: A tool to predict the cross reactivity of peptides complexed with Major Histocompatibility Complex I.Front Immunol. 2022 Oct 28;13:930590. doi: 10.3389/fimmu.2022.930590. eCollection 2022. Front Immunol. 2022. PMID: 36389840 Free PMC article.
-
Mutations in glioblastoma proteins do not disrupt epitope presentation and recognition, maintaining a specific CD8 T cell immune response potential.Sci Rep. 2024 Jul 19;14(1):16721. doi: 10.1038/s41598-024-67099-2. Sci Rep. 2024. PMID: 39030304 Free PMC article.
-
TCRpMHCmodels: Structural modelling of TCR-pMHC class I complexes.Sci Rep. 2019 Oct 10;9(1):14530. doi: 10.1038/s41598-019-50932-4. Sci Rep. 2019. PMID: 31601838 Free PMC article.
References
-
- Dzik JM. The ancestry and cumulative evolution of immune reactions. Acta Biochim Pol (2010) 57(4):443–66. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials