

Reversible differentiation of immortalized human bladder smooth muscle cells accompanied by actin bundle reorganization
- PMID: 29049386
- PMCID: PMC5648286
- DOI: 10.1371/journal.pone.0186584
Reversible differentiation of immortalized human bladder smooth muscle cells accompanied by actin bundle reorganization
Abstract
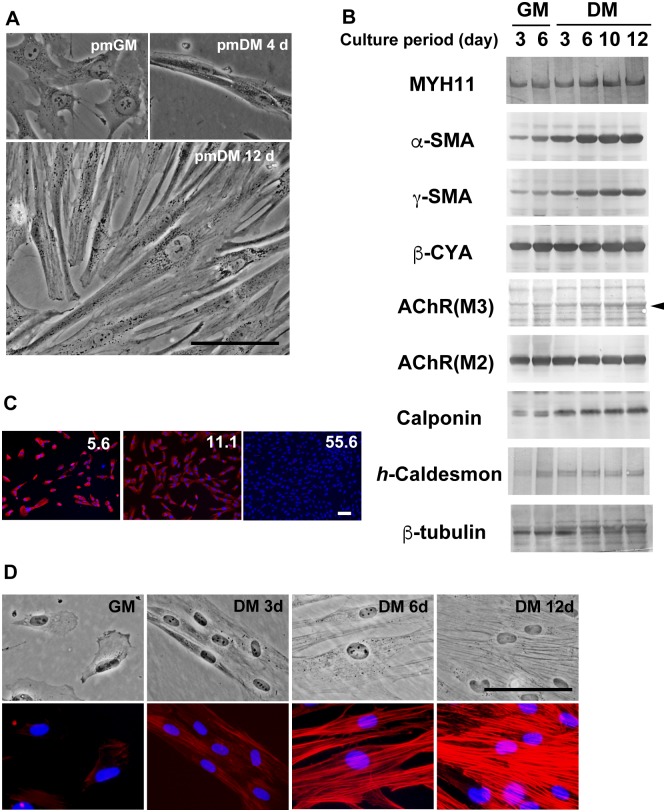
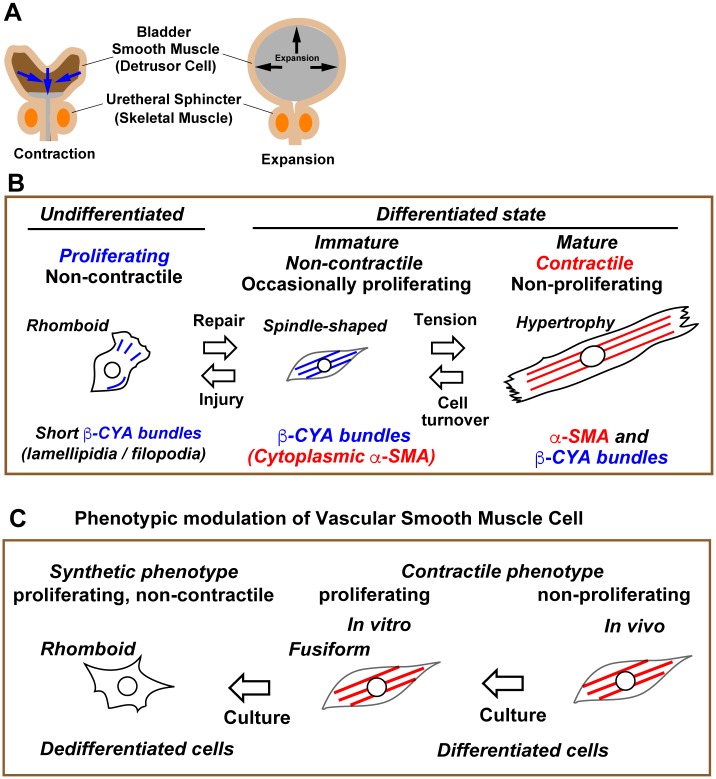
Previous studies have shown that phenotypic modulation of smooth muscle cells (SMCs) plays a pivotal role in human diseases. However, the molecular mechanisms underlying the reversible differentiation of SMCs remain elusive particularly because cultured SMCs that reproducibly exhibit bidirectional phenotypic modulation have not been established. Here we established an immortalized human bladder SMC line designated as hBS11. Under differentiation-inducing conditions, hBS11 cells underwent smooth muscle differentiation accompanied by the robust expression of smooth muscle differentiation markers and isoform-dependent reorganization of actin bundles. The cholinergic receptor agonist carbachol increased intracellular calcium in differentiated hBS11 cells in an acetylcholine muscarinic receptor-dependent manner. Differentiated hBS11 cells displayed contractile properties depending on the elevation in the levels of intracellular calcium. Depolarization of membrane potential triggered inward sodium current in differentiated hBS11 cells. However, differentiated hBS11 cells lost the differentiated phenotype and resumed mitosis when re-fed with growth medium. Our study provides direct evidence pertaining to the human bladder SMCs being able to retain the capacity of reversible differentiation and that the reorganization of actin bundles is involved in the reinstatement of contractility. Moreover, we have established a human SMC line retaining high proliferating potential without compromising differentiation potential.
Conflict of interest statement
Figures






References
-
- Campbell JH, Campbell GR. Smooth muscle phenotypic modulation—a personal experience. Arteriosclerosis, thrombosis, and vascular biology. 2012;32(8):1784–9. doi: 10.1161/ATVBAHA.111.243212 . - DOI - PubMed
-
- Halayko AJ, Solway J. Molecular mechanisms of phenotypic plasticity in smooth muscle cells. Journal of applied physiology. 2001;90(1):358–68. . - PubMed
-
- Rensen SS, Doevendans PA, van Eys GJ. Regulation and characteristics of vascular smooth muscle cell phenotypic diversity. Netherlands heart journal: monthly journal of the Netherlands Society of Cardiology and the Netherlands Heart Foundation. 2007;15(3):100–8. doi: 10.1007/BF03085963 - DOI - PMC - PubMed
-
- Owens GK, Kumar MS, Wamhoff BR. Molecular regulation of vascular smooth muscle cell differentiation in development and disease. Physiol Rev. 2004;84(3):767–801. doi: 10.1152/physrev.00041.2003 . - DOI - PubMed
-
- Xie C, Ritchie RP, Huang H, Zhang J, Chen YE. Smooth muscle cell differentiation in vitro: models and underlying molecular mechanisms. Arteriosclerosis, thrombosis, and vascular biology. 2011;31(7):1485–94. doi: 10.1161/ATVBAHA.110.221101 - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
